ENDOCRINOLOGIA: HORMONAS, RECEPTORES, MECANISMOS DE ACCION HORMONAL.
GENERALIDADES
La endocrinología es un campo de la ciencia que se encarga del estudio de la síntesis, secreción, función hormonal, y de los mecanismos de regulación de la secreción hormonal. Etimológicamente endocrino proviene de dos vocablos griegos, endon: dentro y Krinein: secretar o separar; esto es, secretar al interior. El término Hormona proviene también de un vocablo griego: hormein que significa excitar (1).
El sistema endocrino conjuntamente con el sistema nervioso constituyen dos de los más importantes sistemas que permiten mantener la homeostasis ó medio interno del organismo. Ambos constituyen mecanismos de defensa frente a los cambios ambientales; el sistema nervioso participa en la respuesta inicial frente a un estímulo, pero su acción es de corta duración. Si el estímulo persiste entra en acción el sistema endocrino, cuya respuesta es más tardía pero de mayor duración.
El sistema endocrino tiene un rol importante pues interviene regulando los mecanismos necesarios en los procesos de adaptación de las especies a los cambios ambientales.
Adaptación es la capacidad que tienen los individuos para desarrollar actividad física de diversa intensidad y duración sin que ello afecte su salud, y de poder reproducirse en un medio ambiente diferente al habitual. En la adaptación entran en juego las variaciones genéticas y los factores biológicos (fenotípicos) que permiten a los individuos nacer, crecer y reproducirse en un ambiente diferente al original en forma natural y normal; en estas circunstancias los individuos son capaces de realizar esfuerzos físicos extremos sin demedro de su salud (2). Como ejemplo podemos mencionar a las poblaciones de altura en los Andes que habitan allí desde hace 12,000 años AC.
Muchas especies han desaparecido del planeta porque no fueron capaces de adaptarse a los diversos cambios ambientales, muchos de ellos climáticos, que han ocurrido en el mundo a través del tiempo. Algunas han desaparecido porque perdieron su capacidad de reproducirse, otras porque limitaron su actividad física lo que impedía conseguir su sustento alimenticio, y otras porque no desarrollaron mecanismos que eviten enfermarse.
La vida en las ciudades representa un cambio ambiental de numerosas poblaciones habituadas durante siglos a la vida rural. Durante los últimos 100 años se ha desarrollado una migración masiva en muchas partes del mundo de pobladores habituados al campo a un nuevo ambiente, como consecuencia de la industrialización y de la urbanización. Así, se han desarrollado las ciudades, que sin embargo, ha llevado a que se desarrolle una patología poca vista en los medios rurales, como es la diabetes mellitus. Cambios en los hábitos alimenticios entre otros han permitido que genes de la diabetes que no se expresaban en los pobladores en su ambiente rural, si se expresen en el ambiente urbano.
Otro cambio ambiental, como se ha establecido anteriormente, es el que le ha ocurrido a los seres que viven en las alturas andinas. La población humana americana es originaria de zonas a nivel del mar, que migraron del Asia hacia el continente americano por el estrecho de Berhing, durante la última glaciación. A las zonas andinas peruanas llega hace aproximadamente 20,000 años según Richard McNeish. De acuerdo a Bonavía, la antiguedad del hombre peruano en el ande debe ser considerado en 12,000 años (3). El hombre para vivir en zonas de gran altitud (>3,000 años) debe primero aclimatarse. La aclimatación se presenta en los individuos que están temporalmente expuestos a la altura, y que en cierto grado les permite tolerar la altura. En esta fase hay un incremento en la eritropoyesis (producción de glóbulos rojos), y con ello se incrementa la concentración de hemoglobina en la sangre, y mejora la capacidad de transporte de oxígeno. Cerca del 20% de personas que habitan por encima de 4000 metros hacen una eritrocitosis excesiva (hematocrito >65%) que les impide tolerar la altura y desarrollan el mal de montaña crónico. Estos sujetos deben ser trasladados a zonas de baja altitud para mejorar su estado de salud. Estos sujetos son considerados como desadaptados o no adaptados.
Se cree que los más de 13,000 años que ha residido el hombre (o mujer) en la altura ha sido suficiente para que ellos se adapten, y que la intromisión o mestizaje hispano producto de la conquista española en el siglo XVI haya detenido o revertido parcialmente el proceso de adaptación. Ello explicaría porque en la actualidad se puede apreciar personas que viven normalmente en zonas por encima de los 3,000 metros de altura, en tanto que otras no lograr sobrellevar estas alturas.
En resumen la adaptación es un proceso que se desarrolla en muchas generaciones y donde interviene fundamentalmente el sistema endocrino, donde se realizan una serie de cambios que apuntan a mantener la homeostasis del organismo y la capacidad reproductiva. El mantenimiento de la homeostasis implica en desarrollar mecanismos adecuados para regular la energía del metabolismo.
De lo anteriormente expuesto podemos decir que la endocrinología puede ser dividida en dos grandes áreas de estudio:
a.- El metabólico.
b.- El reproductivo
Otras funciones del sistema endocrino son aquellas referidas a los procesos de diferenciación celular, y en el control del sistema inmunológico.
En el primer caso tenemos como ejemplo el rol de las hormonas tiroideas en el proceso de la mielinización de las fibras nerviosas; así, su deficiencia (hipotiroidismo) congénita resulta en cretinismo, una patología donde se observa retardo mental. Igualmente los andrógenos gonadales son importantes para la diferenciación de los genitales externos, los conductos reproductivos internos, y del cerebro masculino.
En el segundo caso existe un sistema de regulación a través de los glucocorticoides que modulan la respuesta inmune; en éste sistema también participa el sistema nervioso, por lo que podemos calificarlo como sistema neuro-inmuno-endocrino.
Existen hormonas que tienen tanto función metabólica como reproductiva. Entre ellas tenemos a la testosterona, el estradiol y la leptina.
Endocrinología y Metabolismo.
El sistema endocrino regula el metabolismo:
1. hidro-electrolítico
2. del calcio y del fósforo (sistema óseo)
3. de los carbohidratos
4. de los lípidos y
5. de las proteínas.
La regulación del anabolismo (producción) y catabolismo (degradación) de estos compuestos, está orientada a regular la energía necesaria para las actividades celulares, el crecimiento y el desarrollo. En situaciones negativas como por ejemplo desnutrición, el organismo deja de crecer y desarrollarse para mantener las actividades celulares.
La hormona antidiurética, la hormona atrial natriurética, la aldosterona, la angiotensina, y la prolactina, entre otras, regulan el equilibrio hidrosalino.
La paratohormona, la tirocalcitonina, la vitamina D, el estradiol, la dehidroepiandrosterona, entre otras, son hormonas que regulan el sistema esquelético, y el metabolismo del calcio y fósforo del organismo.
Los glucocorticoides, la insulina, la hormona del crecimiento, las hormonas tiroideas, la somatostatina, y el glucagon, entre otros regulan el metabolismo de los carbohidratos.
La hormona del crecimiento, la leptina, los estrógenos, las hormonas tiroideas, la insulina regulan entre otros, el metabolismo de los lípidos.
Los andrógenos, la insulina, los glucocorticoides, las hormonas tiroideas, la hormona del crecimiento entre otras regulan el metabolismo de las proteínas.
De lo anterior se deduce que hay hormonas que tienen más de una función. Así vemos que las hormonas tiroideas, por ejemplo, regulan el metabolismo de los carbohidratos, de los lípidos y de las proteínas.
Endocrinología y Reproducción.
El sistema endocrino participa de manera fundamental en la regulación de los procesos que permiten el crecimiento y desarrollo de los gametos masculino y femenino. Esto basado a la integración del sistema de regulación: Sistema Nervioso Central - Hipotálamo - Hipófiso - Gonadal. Una alteración en cualquiera de estos niveles de regulación alterará la reproducción y conducirá a la infertilidad.
El sistema nervioso central puede ser regulado exógenamente por estímulos ambientales, que van a influenciar en la disponibilidad de los neurotransmisores.
El sistema endocrino es también importante porque regula el deseo sexual o libido, de regulación tónica en el sexo masculino y de regulación cíclica en el sexo femenino. Esto se puede apreciar claramente en los mamíferos no humanos. Los machos siempre intentan montar a la hembra (regulación tónica) en tanto que la hembra sólo aceptará al macho si está en celo o estro (regulación cíclica). Esta aceptación de la hembra a la monta está asociada al momento en que hay posibilidad de fertilizar (ovulación).
Un ejemplo de como un cambio ambiental afecta la reproducción es la que ocurrió con la conquista española en 1535. La capital del Perú inicialmente localizada en las alturas de Jauja fue trasladada a Lima, entre otras razones porque los caballos y los cerdos no se reproducían en la altura (4), tal como figura en las Actas de Fundación de la ciudad de Lima. Es de entender la importancia que representaban los caballos para los españoles y para la conquista.
SISTEMA ENDOCRINO
El sistema endocrino se fundamenta anatómicamente en las glándulas de secreción interna o glándulas endocrinas. En la definición clásica las glándulas endocrinas típicas están conformadas por células acinares en contacto con una red de vasos sanguíneos. Entre estas tenemos a la hipófisis (pituitaria), tiroides, paratiroides, páncreas, corteza adrenal, y las gónadas. Esta descripción anatómica se ha modificado en los últimos años para dar paso a una clasificación funcional (1), de tal manera que ahora se considera como célula endocrina a toda aquella que secreta una hormona.
En esta clasificación moderna se incluyen al hipotálamo, conformado por neuronas y que sintetizan y secretan a las hormonas liberadoras (GnRH, TRH, CRH, GHRH.), o las hormonas inhibidoras (Somatostatina, Dopamina); al corazón que sintetiza y secreta la hormona atrial natriurética; al pulmón que secreta serotonina y endorfina; al riñón que produce eritropoyetina, y renina; al hígado que sintetiza el factor de crecimiento similar a insulina (IGF) y también a la eritropoyetina; Y al tejido adiposo que produce leptina y también secreta estrona.
El sistema endocrino mantiene una estrecha relación con el sistema nervioso a través del hipotálamo. El hipotálamo anatómicamente es parte del sistema nervioso central, pero como funcionalmente se comporta como sistema endocrino, se le estudia de manera separada y constituye parte del sistema neuroendocrino. El sistema nervioso y el sistema endocrino regulan e integran los procesos fisiológicos permitiendo el funcionamiento armónico de los órganos de los individuos.
La actividad del sistema endocrino es ejecutada por las hormonas mientras que la del sistema nervioso central la es por los neurotransmisores. El lugar de acción de un neurotransmisor o de una hormona se denomina órgano blanco o diana. La forma de acción en el órgano blanco es directa en el sistema nervioso a través del espacio intersináptico, e indirecta en el sistema endocrino a través de la vía sanguínea.
Hay sustancias que pueden actuar tanto como neurotransmisores y como hormonas, dependiendo su denominación del tipo de acción que realiza. Por ejemplo, si la serotonina es liberada en los núcleos del rafe dorsal (tallo cerebral) al hipocampo actuará como neurotransmisor, pero si es liberada a la circulación y actúa por vía sanguínea actúa como hormona (5).
TIPOS DE TRASMISION QUIMICA
Las células interactúan unas a otras y modifican su actividad metabólica por acción de sustancias químicas denominadas transmisores. Existen diferentes tipos de transmisión química: neurocrina, neuroendocrina, endocrina, paracrina y autocrina (Figura 1).
Transmisión Neurocrina
Es la que ocurre en el sistema nervioso a través de la liberación de sustancias químicas denominadas neurotransmisores, al espacio intersináptico y que se une a un receptor post-sináptico modificando la actividad metabólica de la célula post-sináptica.
Transmisión Endocrina.
Es la que ocurre en el sistema endocrino a través de la liberación de sustancias químicas denominadas hormonas, que actúan a distancia sobre una célula efectora.
Transmisión Neuroendocrina
Es la que ocurre por la liberación de sustancias químicas (neurohormonas) en los terminales nerviosos hacia la circulación, y actúan a distancia sobre una célula efectora. El ejemplo clásico es la secreción de las neurohormonas (hormonas liberadoras e inhibidoras) del hipotálamo a la eminencia media y que a través de la vía sanguínea porta-hipofisiaria se van a trasladar a la adenohipófisis (hipófisis anterior) donde van a actuar.
Transmisión paracrina.
Es la transmisión que ocurre entre dos células adyacentes, donde una de las células secreta la sustancia (parahormona), que actúa por difusión en la célula vecina modificando su función. Se le conoce también como control local. En este caso no hay participación de la vía sanguínea. Bajo este sistema de transmisión se puede regular la acción de una hormona aumentando o disminuyendo su acción.
Transmisión autocrina
Es cuando una sustancia química actúa sobre la misma célula que la produce para regular su secreción.

Figura 1. Representación esquemática de la transmisión química
HORMONAS
En 1902, Bayliss y Starling definieron por primera vez el concepto de hormona, al descubrir la secretina, liberada a la circulación por la mucosa duodenal, y que estimula el flujo pancreático. Estos autores definieron a la hormona como cualquier sustancia producida normalmente en las células de una región del organismo y llevada por el torrente sanguíneo a otras partes, sobre las cuales actúa con efecto favorable para el organismo en conjunto.
Actualmente se definen a las hormonas como compuestos químicos secretados en mínimas concentraciones al torrente sanguíneo por células específicas (pueden ser glándulas endocrinas clásicas o no), y que actúan en células distantes al lugar de origen, donde se unen a receptores específicos produciendo una respuesta biológica.
Existen sustancias que simulan el efecto de una hormona pero no son hormonas; éstas son la glucosa, los ácidos grasos no esterificados, y las prostaglandinas. La glucosa actúa sobre el páncreas, en un receptor específico y libera insulina, pero no es hormona porque actúa en altas concentraciones (miligramos), en tanto que las hormonas actúan a mínimas concentraciones: picogramos (10-12 g) y nanogramos (10-9 g). La misma situación ocurre para los ácidos grasos no esterificados, cuya liberación produce inhibición de la secreción de hormona del crecimiento por la adenohipófisis. Las prostaglandinas simulan la función hormonal, pero se producen localmente, en el lado interno de la membrana celular, y no actúan fisiológicamente a distancia. Farmacológicamente es posible administrar prostaglandinas y tener un efecto a distancia.
Muchas de las hormonas secretadas por las células endocrinas son inactivas (precursoras) y requieren transformarse en otra molécula para tener actividad biológica. Por ejemplo, la tiroxina (T4) secretada por la glándula tiroides requiere perder un iodo y transformarse en tri-iodotironina (T3) para ser biológicamente activa. En este caso para ser activa se ha eliminado un yodo. Igualmente, la testosterona debe transformarse por acción de una enzima la 5 alfa reductasa en dihidrotestosterona en muchos tejidos para ejercer su acción. En este caso ha ocurrido una reducción (incorporación de un hidrógeno) para ser biológicamente activa.
HORMONAS Y PROTEINAS LIGADORAS
Las hormonas esteroidales (andrógenos, estrógenos, progesterona, corticoides) y las tiroideas circulan en la sangre tanto ligada a una proteína como en forma libre, siendo ésta última la biológicamente activa. Generalmente las determinaciones hormonales se refieren a la concentración total (hormona libre + hormona ligada a la proteína), y no siempre una alteración en los niveles de la hormona total refleja una alteración de la fracción libre, puesto que existen muchas situaciones en que se afecta el contenido de la proteína ligadora sin que necesariamente ocurra una disfunción hormonal. Un ejemplo, es el incremento de la globulina ligadora de tiroxina (TBG) por acción de los estrógenos incrementados durante el embarazo; la tiroxina total se incrementa pero no la fracción libre, por lo tanto no hay hipertiroidismo.
La globulina ligadora de hormonas sexuales (SHBG) también se incrementa por acción de los estrógenos y progestágenos. Estas proteínas se unen a los estrógenos, andrógenos y progestágenos.
Si bien las globulinas ligadoras no parecen tener otra función que la de determinar la fracción ligada de la hormona (no activa), recientemente se ha demostrado que la globulina ligadora de hormonas sexuales después de ligarse a la hormona, también puede unirse a un receptor específico en la membrana celular y consecuentemente activar AMPc y proteína kinasa A (6) y por ende tener una acción hormonal.
Existen hormonas proteicas como la hormona de crecimiento que se unen a proteínas ligadoras circulantes en sangre (7,8) como paso necesario para su acción hormonal. Se considera a esta proteína ligadora como el receptor extracelular, pues su ausencia resulta en falla de la acción de la hormona del crecimiento. Esto es, para la acción hormonal, la hormona de Crecimiento (GH) se une al receptor celular, y este complejo hormona-receptor extracelular (proteína ligadora) se une a un receptor de membrana, y a partir de allí se produce la cascada que detrrmina finalmente la acción de la hormona, en este caso la hormona de crecimiento..
En el humano, se produce esta proteína ligadora de hormona de crecimiento por la ruptura enzimática de la porción extracelular del receptor de membrana en el hígado (7).Esto también se ha observado para la acción de ciertas hormonas esteroidales que se unen a la SHBG y el complejo se une a un receptor de membrana; así, el estradiol se liga a la SHBG, y el complejo estradiol-SHBG se une a un receptor de membrana en las células prostáticas estimulando la generación de AMPc y posteriormente la activación del receptor de andrógenos (9).
RECEPTORES
Muchas células son expuestas a las hormonas, sin embargo, sólo algunas responden. De esto nace el concepto de especificidad. Tal especificidad de la acción hormonal parece residir en la presencia de receptores en el órgano blanco que pueden reconocer específicamente su señal. Se ha comparado este proceso, como aquel que ocurre entre la llave y su cerradura.
Los receptores son proteínas cuyo número y afinidad pueden modificarse de acuerdo a las circunstancias. Estos pueden ser de membrana, citoplásmicas y nucleares. Por lo general tienen receptores de membrana aquellas hormonas que por su tamaño no pueden entrar a la célula o aquellas que por su poca liposolubilidad tampoco lo pueden hacer. Las proteínas no pueden atravesar la membrana por su tamaño, en tanto que los esteroides que son moléculas pequeñas y liposolubles si la atraviesan.
Las hormonas amínicas (serotonina, dopamina), peptídicas (GnRH) y las proteicas se unen a receptores de membrana, y las hormonas esteroidales lo hacen a receptores intracelulares. Recientemente se ha demostrado que algunos sistemas celulares tienen receptores de membrana para los esteroides; por ejemplo el espermatozoide tiene receptores de membrana para progesterona. Los receptores de los andrógenos pertenecen a una superfamilia de receptores nucleares que emplean mecanismos genéticos complejos para controlar el desarrollo y las funciones de los tejidos “blanco”. Los receptores de andrógenos activan o reprimen la transcripción de genes a través de su asociación a segmentos específicos del DNA (Elemento de Respuesta) y/o proteínas (10).
Las hormonas que actúan a través de receptores de membrana activan la formación de un segundo mensajero, que lleva el mensaje de la hormona al interior de la célula para la elaboración de la respuesta biológica o acción hormonal. En la membrana plasmática existe también un tipo de receptores que son tirosinas kinasas. En este tipo de receptores se unen la insulina, el factor de crecimiento similar a insulina (IGF), y el factor de crecimiento epidermal (EGF). De lo anterior se deduce que la hormona se constituye en el primer mensajero.
En algunos casos es necesario que la hormona se una previamente a una proteína ligadora que circula en sangre y sólo después de esta unión se puede activar el receptor de membrana. En estos casos se considera a la proteína ligadora como el receptor extracelular. Este es el caso para la proteína ligadora de hormona del crecimiento (8), y en algunos casos para la globulina ligadora de hormonas sexuales (SHBG) (9).
MECANISMO DE ACCION HORMONAL
La respuesta de un organismo a la administración de una hormona puede ser considerada bajo tres modalidades: función, mecanismo de acción, y el efecto biológico.
1. La función se refiere al propósito o utilidad de la hormona respecto a la regulación metabólica o a los cambios metabólicos que produce.
2. El mecanismo de acción se refiere a como una hormona interactúa con un receptor específico y todos los eventos intracelulares subsiguientes que conllevarán al efecto biológico.
3. El efecto biológico es la respuesta medible que produce la hormona sobre un órgano o acción enzimática.
Las hormonas influencian los estados funcionales y morfogenéticos de tejidos que se encuentran distantes de las glándulas endocrinas que las producen. Otras, como los andrógenos participan en la diferenciación celular, la proliferación así como en la carcinogénesis (vg. cáncer de próstata) (10). El sistema endocrino también regula el sistema inmunológico. Por ejemplo, los cambios cíclicos en la respuesta inmune tienen implicancia fisiológica, tal como la disminución o supresión de la inmunidad mediada por células, para evitar que los espermatozoides, y el embrión pre-implantacional sean reconocidos como extraños (11), y de esta manera favorecer la fertilización y la implantación respectivamente.
Mecanismo de acción para hormonas con receptores de membrana.
Las hormonas con receptores de membrana actúan produciendo a nivel intracelular sustancias denominadas “segundo mensajeros”. Un segundo mensajero es una sustancia cuya concentración aumenta intracelularmente en respuesta a la hormona primaria (primer mensajero). Su función es la de llevar la señal hormonal al interior de la célula, con la finalidad de traducirla en acción biológica.
Entre los segundos mensajeros tenemos: el AMP cíclico, el GMP cíclico, el ión calcio, el ión calcio unido a la calmodulina, el ácido araquidónico y sus metabolitos, el inositol trifosfato, y el diacilglicerol.
Los segundos mensajeros actúan fosforilando proteínas que a su vez van a actuar sobre porciones específicas del DNA denominadas elementos de respuestos, y a partir de allí ejercer la acción hormonal.
AMP cíclico
El descubrimiento del AMPc permitió a Sutherland obtener el premio Nóbel en Medicina en 1958, y su participación en los procesos hormonales fue ampliamente reconocida en la década de los sesenta.
El AMPc es producido por la adenil ciclasa que es una enzima unida al lado interno de la membrana citoplasmática. Esta enzima convierte ATP en AMP cíclico en presencia de iones Mg2+. La concentración de AMPc en el citoplasma es a su vez controlada por una segunda enzima, la fosfodiesterasa que cataliza la hidrólisis de AMPc en 5’-AMP.
El AMPc activa una proteína kinasa. La proteína kinasa nativa está compuesta de dos subunidades: catalítica y regulatoria. La subunidad regulatoria suprime la actividad de la proteína kinasa nativa mediante su asociación con la subunidad catalítica. La activación de la proteína kinasa ocurre después que se ha liberado la subunidad catalítica de la subunidad regulatoria, por la unión del AMPc a la subunidad regulatoria de la proteína kinasa.
La proteína kinasa activa (subunidad catalítica) fosforila proteínas específicas denominadas:
1. Proteína ligadora del elemento de respuesta al AMPc (CREB)
2. Modulador del elemento de respuesta al AMPc (CREM).
La fosforilación de estas proteínas produce cambios en sus actividades lo que les permiten interactuar con secuencias específicas del DNA denominadas ELEMENTOS DE RESPUESTA AL AMPc (CRE) para modular la transcripción del gen. Tanto la CREB como el CREM pertenecen a una familia de proteínas ligadoras que tienen alguna homología entre sí. Esta familia de proteínas ligadoras incluyen tanto inhibidores como activadores de la transcripción de los genes.
En los efectos tróficos de respuesta prolongada, el sistema regula la síntesis proteica a nivel de la transcripción y la translación de los ácidos nucleicos.
Las gonadotropinas en las gónadas son buen ejemplo de la activación del AMPc.
GMP cíclico
El GMPc es producido por acción de la enzima guanidil ciclasa que convierte GTP a GMPc. La GMPc es posteriormente inactivada por acción de una fosfosdiesterasa. La GMPc activa la proteína kinasa G que a su vez fosforila proteínas.
La hormona atrial natriurética y el óxido nítrico actúan a través de la activación del GMPc.
FOSFODIESTERASAS
Las fosfodiesterasas son enzimas que permiten la regulación de la concentración intracelular de AMPc y GMPc. En el organismo se encuentran 5 tipos de fosfodiesterasas:
1. Fosfodiesterasa I: Estimulada por el complejo calcio-calmodulina
2. Fosfodiesterasa II: Estimulada por GMPc
3. Fosfodiesterasa III: Inhibida por GMPc
4. Fosfodiesterasa IV: Específico para AMPc
5. Fosfodiesterasa V: Específico para GMPc.
Por ejemplo, las fosfodiesterasas III, IV y V se encuentran en el tejido cavernoso del pene, y regulan la erección. El sildenafil (viagra) actúa inhibiendo la fosfodiesterasa V y favorece la erección del pene (12).
CALCIO
En la década de los setenta empieza a tomar impulso el rol del calcio en los procesos endocrinos, particularmente en el acoplamiento estímulo-secreción.
El calcio actúa como segundo mensajero al incrementarse como fracción libre intracitoplasmática o al unirse a la calmodulina. En este modelo, la hormona específica se une a un receptor en el lado externo de la membrana; esto conduce a un cambio conformacional en la membrana que directamente estimulará la apertura o la constitución de los canales de calcio que favorecerán el influjo de calcio.
Los iones de calcio pueden afectar los procesos celulares directamente o pueden ligarse a la calmodulina. El complejo calcio-calmodulina puede ligarse a ciertas enzimas con la finalidad de activarlos. Este complejo activa una proteina kinasa en el núcleo, fosforilándolo y activando a la proteína ligadora dependiente de AMPc (CREB y el elemento de respuesta dependiente de AMPc (CRE) (13,14). La proteína kinasa nuclear dependiente de calcio-calmodulina puede funcionar como un mediador de la transcripción inducida por calcio (14,15) en sistemas endocrinos y neuroendocrinos como el hipocampo, hipotálamo y pituitaria (14,16).
El calcio intracelular puede activar los sistemas de secreción formando un puente de los gránulos de secreción con la membrana plasmática desarrollando el acoplamiento estímulo-secreción. Otra forma de aumentar el calcio intracelular es a través de su liberación de los compartimentos celulares, tales como el retículo endoplásmico y la mitocondria. Este proceso puede ser desarrollado por acción de un segundo mensajero, el inositol trifosfato (ITP), que a su vez procede de la hidrólisis del fosfatidil-inositol-4,5-bifosfato.
El calcio intracelular puede participar en los procesos de exocitosis para la secreción hormonal, o activar sistemas enzimáticos que participan en la formación de otros mensajeros.
FOSFOINOSITOLES
El conocimiento de que el metabolismo de los fosfolípidos de membrana intervienen como reguladores de la acción hormonal aparece en la década de los ochenta.
Los fosfoinositoles (fosfatidilinositol, fosfatidil inositol monofosfato, y fosfatidil inositol bifosfato) se encuentran en la cara interna de la membrana citoplasmática. De todos estos, el fosfatidilinositol-4,5-bifosfato es el que interviene después que la hormona se une a su receptor. Según el modelo, la hidrólisis de este fosfolípido por acción de una fosfodiesterasa produce diacilglicerol (DAG) e inositol-1,4,5-trifosfato (ITP).
El diacilglicerol siempre tiene en el carbono 1 el ácido graso: Acido araquidónico.
El ITP actúa movilizando calcio intracelular, mientras que el diacilglicerol puede activar una proteína kinasa C (PKC) o liberar ácido araquidónico.
La activación de una proteína kinasa C por acción de la DAG, induce la fosforilación de proteínas que favorecen la respuesta biológica u hormonal. El DAG por acción de una lipasa dependiente de calcio puede liberar ácido araquidónico de la posición 1 del DAG. El ácido araquidónico puede activar la formación de GTP a GMP cíclico, o ser metabolizada por la enzima lipo-oxigenasa para producir hidroxiperóxidos y leucotrienos, o por la enzima ciclo-oxigenasa para producir prostaglandinas, tromboxano, y prostaciclina. Varios de estos metabolitos actúan como segundo mensajeros.
Las hormonas pueden actuar en cualquiera de estas vías; así, la insulina, la angiotensina II y las hormonas hipotalámicas actúan a través de la proteína kinasa C, el EGF a través de la vía de la lipo-oxigenasa, la serotonina a través de la vía de la ciclo-oxigenasa; la hormona atrial natriurética activa el sistema guanilato ciclasa-GMPc.
Mecanismo de acción para hormonas con receptores intracelulares
A diferencia de las hormonas peptídicas, que debido a su peso molecular no pueden penetrar a la célula, los esteroides y las hormonas tiroideas, por su bajo peso molecular y por su naturaleza lipofílica atraviezan con facilidad la membrana citoplasmática. Aunque los esteroides y las hormonas tiroideas penetran a todas las células del organismo, sólo aquellas células que contienen receptores específicos para ellas responderán al estímulo hormonal.
Los esteroides son transportados en el torrente sanguíneo en forma libre o ligados a proteínas séricas, como la globulina ligadora de hormonas sexuales (SHBG), la globulina ligadora de corticosteroides (CBG), y la albúmina.
En las células de los órganos blanco, los esteroides ingresan por difusión, aunque como ya se ha dicho anteriormente, se puede unir a la SHBG en el suero y unirse a un receptor de membrana y activar la formación de AMPc. Si el esteroide difunde dentro de la célula por difusión, permanece dentro de ella por un tiempo largo, por lo que puede mantenerse una concentración intracelular aumentada, a pesar de que los niveles plasmáticos vayan disminuyendo. La razón de esta diferencia se explica por la presencia en el núcleo de las células efectoras de una proteína de alto peso molecular y con una alta afinidad para ligar selectivamente un esteroide dado, y que es conocido como receptor.
El receptor se ubica dentro del núcleo, y la unión del esteroide al receptor induce a un cambio conformacional del receptor que mejora su afinidad para secuencias específicas en el DNA denominadas Elementos de Respuesta a los Esteroides (ERE). Esta reacción, a su vez induce cambios en la expresión de los genes que finalmente genera la síntesis de proteína y la respuesta celular.
La asociación de esteroide al receptor precede cualquier efecto fisiológico, más aún, los tejidos que carecen del receptor, no responderán al estímulo hormonal. Este es el caso del testículo femenizante donde el testículo fetal produce testosterona pero al no haber receptores para testosterona, no se produce la acción de esta hormona.
El receptor de andrógenos pertenece a una familia de proteínas que actúan como reguladores de la transcripción de genes por ligazón a secuencias específicas del DNA (17). Esta familia incluye a los receptores de las hormonas tiroideas, vitamina D y ácido retinoico. La existencia de esta gran familia de proteínas nucleares sugiere que la acción de los esteroides en el genoma puede representar un mecanismo de control de la transcripción específico que es utilizado por la célula para diferentes propósitos además de mediar la respuesta hormonal.
Los receptores nucleares han sido clasificados en dos categorías:
Grupo I, que comprende a los receptores para estrógenos, hormonas tiroideas, ácido retinoico, y vitamina D, y que reconocen los flancos 5’ y 3’ de los elementos de respuesta del DNA.
Grupo II, que comprende a los receptores para andrógenos, progestágenos, glucocorticoides y mineralocorticoides, que reconoce el flanco 5’ del DNA.
La secuencia de bases en la mitad 3’ del DNA parece importante pues la alteración de 1 ó 2 bases dentro de este sitio es suficiente para convertir un elemento de respuesta al estrógeno en un elemento de respuesta del grupo II.
La mayoría de interacciones esteroide-receptor ocurre dentro del núcleo, excepto el receptor de los glucocorticoides, que parece en alguna extensión translocarse del citoplasma al núcleo después de haberse ligado a la hormona.
La activación de los elementos de respuesta induce que la enzima RNA polimerasa ocupe el sitio de iniciación para que sintetice el RNAm. El RNA mensajero migra hacia los ribosomas del citoplasma donde se sintetiza la proteína cuya codificación ha sido desreprimida.
En conclusión, los esteroides modulan la transcripción de genes por su interacción con secuencias específicas de genes en el DNA. Estas secuencias son conocidas como elementos de respuesta, y facilitan que sintetice RNA mensajero durante la transcripción, seguido por la síntesis proteica durante la translación, y en esta forma producir los efectos fisiológicos (18).
SISTEMA DE REGULACION HORMONAL
La secreción hormonal es regulada por estímulos directos y por mecanismos de retroalimentación. Los sistemas hormonales se integran en ejes donde hay un sistema de regulación superior, conformado por el sistema nervioso central (SNC), que a través de una regulación neurocrina actúa sobre el hipotalálamo. El hipotálamo es la glándula maestra a partir del cual se desarrolla la integración con la hipófisis. Esto quiere decir que el SNC, el hipotálamo y la hipófisis son comunes para todos los ejes de regulación hormonal; a partir de la hipófisis se diversifican las funciones.
Así tenemos, el eje SNC-hipotálamo-hipófiso-gonadal; eje SNC-hipotálamo-hipófiso-tiroideo; eje SNC-hipotálamo-hipófiso-córtico adrenal; eje SNC-hipotálamo-hipófiso-pancreático entre otros.
REGULACION DIRECTA
La regulación directa es la que ocurre de un nivel superior a otra de nivel inferior. En la Figura 2 se observa que la glándula A regula directamente la secreción de la glándula B; por ejemplo, la hipófisis (Glándula A) que secreta la hormona luteinizante (LH) estimula la secreción de la testosterona por las células de Leydig en el testículo (Glándula B).
RETROALIMENTACION
La retroalimentación es la regulación a partir de una glándula del nivel inferior hacia la glándula que la estimula y que está en un nivel superior. Este sistema permite mantener el equilibrio en la secreción hormonal para evitar que una glándula de nivel inferior se mantenga sobre-estimulada por una glándula de nivel superior. Por ejemplo, la hipófisis (nivel superior) secreta hormona del crecimiento (GH), y ésta actúa sobre el hígado produciendo, el factor de crecimiento similar a insulina-I ó IGF-I (nivel inferior), que a su vez actúa sobre el crecimiento de los huesos. Si no ocurriera retroalimentación, la GH seguiría actuando y se produciría crecimiento desmesurado del hueso (gigantismo). La IGF-I por retroalimentación inhibe la secreción de GH.

Retroalimentación negativa
La retroalimentación es negativa cuando el producto final (B en la figura 2b) inhibe la secreción de la glándula A. Por ejemplo, la hormona luteinizante (LH) estimula en las células de Leydig del testículo la producción de testosterona, la cual al circular por la sangre llega a la hipófisis donde va a inhibir la secreción de LH.
Retroalimentación positiva
La retroalimentación es positiva cuando la secreción de una célula efectora (B en la figura 2a) incrementa la secreción de la hormona que la estimula (A en la figura 2a); éste mecanismo de regulación es el que ocurre menos frecuentemente en el organismo. Un ejemplo es la liberación cíclica de gonadotropinas en la fase pre-ovulatoria de la mujer. El producto (estradiol) de la célula efectora (célula ovárica), que es a su vez estimulada por la hormona luteinizante (LH) y la folículo estimulante (FSH), van a determinar una mayor secreción de LH y FSH por la hipófisis (pico ovulatorio de LH y FSH).
CLASIFICACION DE LAS HORMONAS
Las hormonas según su estructura química pueden ser aminas, péptidos, proteínas o esteroides.
1. Aminas
- Hipotalámica: Dopamina
- Tiroideas: Tri-iodotironina (T3) y Tiroxina (T4)
- Médula suprarrenal: Epinefrina
2. Péptidos
Están conformados por menos de 100 aminoácidos unidos por enlaces peptidérgicos
- Hormonas hipotalámicas: Hormona liberadora de corticotrofina (CRH), Hormona liberadora de hormona del crecimiento (GHRH), Hormona liberadora de gonadotropinas (GnRH), Hormona liberadora de tirotropina (TRH), somatostatina.
- Hormonas hipofisiarias: Corticotrofina (ACTH), ß-endorfinas, hormona antidiurética (ADH), oxitocina.
- Hormonas pancreáticas: Glucagon, insulina, somatostatina.
- Hormonas reguladoras del calcio: tirocalcitonina, paratohormona.
- Hormona del corazón: hormona atrial natriurética.
- Hormona de las células endoteliales: endotelinas.
3. Proteínas. Están conformadas por más de 100 aminoácidos sólos (proteicas) o ligados a carbohidratos (Glucoproteicas).
Hormonas proteicas
- Hormonas hipofisiarias: Tirotropina (TSH), Hormona del crecimiento, Prolactina.
Hormonas glicoproteicas
- Hormonas hipofisiarias: Hormona luteinizante (LH), Hormona folículo estimulante (FSH), y TSH.
- Hormonas placentarias: Hormona coriónica gonadotropa (hCG), y Tirotropina placentaria
- Hormonas gonadales: Inhibina, y Activina.
4. Esteroides
Las hormonas esteroidales son lipofílicas y se producen en glándulas cuyo origen embriológico es mesodérmico, tales como la corteza adrenal, el ovario y el testículo. El núcleo fundamental de estas hormonas es el ciclopentanoperhidrofenantreno (Figura 3 ejemplos).
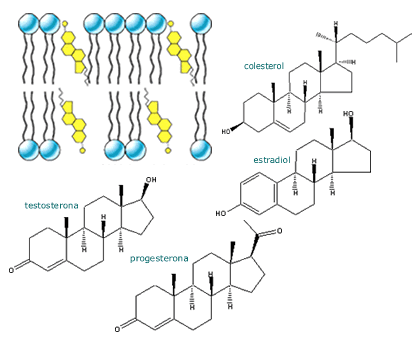
Las hormonas esteroidales según el número de átomos de carbono pueden dividirse en pregnanos (21 carbonos), androstanos (19 carbonos) y estranos (18 carbonos). Los ejemplos típicos de estos grupos son la progesterona, la testosterona y el estradiol, respectivamente.
- Hormonas de la corteza adrenal
Aldosterona
Cortisol y corticosterona
Dehidroepiandrosterona
Dehidroepiandrosterona sulfato
Androstenediona
- Hormonas ováricas
Estrógenos (estrona, estradiol y estriol)
Progesterona
- Hormonas testiculares
Testosterona
Dihidrotestosterona
Estradiol.
HORMONAS Y APOPTOSIS
La proliferación de las células y la muerte celular participan de manera armónica en el mantenimiento de la homeostasis celular (19).
La apoptosis es la muerte celular programada que ocurre normalmente en diferentes estadíos de la morfogénesis, crecimiento y desarrollo de los metazoarios, y en el recambio normal del tejido adulto. Durante la apoptosis, una célula a través de un proceso dependiente de energía ingresa a un suicidio celular iniciado por señales específicas en un microambiente aparentemente normal. La apoptosis se inicia en células específicas por agentes endógenos (hormonas) y exógenos (radiación, químicos, virus) (20).
Las hormonas pueden participar estimulando la apoptosis o previniéndola. Los glucocorticoides estimulan la apoptosis induciendo la muerte de timocitos inmaduros (20,21). La melatonina tiene un efecto preventivo de la apoptosis en las células neuronales inducidas por 6-hidroxidopamina (22).
La Bcl-2 es una proteína que inhibe la apoptosis (23). El estradiol regula la producción de Bcl-2 en varios tejidos, entre ellas, las neuronas del hipotálamo (24). La progesterona administrada simultáneamente con el estradiol, reduce el efecto del estradiol sobre esta actividad protectora. Estos hallazgos sugieren que las hormonas ováricas regulan la Bcl-2 en las neuronas hipotalámicas y que esta proteína puede estar envuelta en los efectos neuroprotectores de los estrógenos.
Una vez iniciada la apoptosis ocurre una cascada de eventos bioquímicos y morfológicos que resultan en una degradación irreversible del DNA genómico y la fragmentación de la célula. La secuencia de eventos de la apoptosis son: agregación de la cromatina, condensación nuclear y citoplasmática, y una eventual fragmentación de la célula en un conglomerado de segmentos con membranas (cuerpos apoptóticos) que a menudo contienen organelas intactas. Estos cuerpos apoptóticos son fagocitadas por los macrófagos. La fragmentación del DNA ocurre antes de que hayan cambios en la permeabilidad de la membrana plasmática y de las intracitoplasmáticas.
REFERENCIAS
1. Gonzales GF. Fisiología Endocrina y de la Reproducción. Rev. Per. Endocr. Metab. 1997;3: 3-26.
2. Gonzales GF y Villena A. Aclimatación y adaptación. En: El futbol y la aclimatación a la altura. Lima:Ediciones IIA. 1998; 23-46.
3. Bonavía D. Perú. Hombre e Historia. De los orígenes al Siglo XV. Tomo I. Lima:Edubanco. 1a Edición. 586 pp, 1991.
4. Gonzales GF. Contribución peruana al estudio de la reproducción humana en la altura, desde los cronistas de la conquista a la actualidad. Acta Andina 7: (en prensa), 1998.
5. Gonzales GF. Serotonina sanguínea: Regulación y significación endocrina. Rev. Soc Per. Endocrinol 1997;3:77-94.
6. Fortunati N, Raineri M, Cignetti A, Hammond GL, Frairia R. Control of membrane sex hormone-binding globulin-receptor (SHBG-R) in MCF-7 cells: effect of locally produced SHBG. Steroids 1998; 63:282-284.
7. Rosenfeld RG. Circulating growth hormone binding proteins. Horm Res 1994; 42:129-132.
8. Fontoura M, Hocquette JF, Clot JP, Tar A, Brauner R, Rappaport R, Postel-Vinay MC. Regulation of the growth hormone binding proteins in human plasma. Acta Endocrinol (Copenh) 1991;124 Suppl 2:10-13.
9. Ding VD, Moller DE, Feeney WP, Didolkar V, Nakhla AM, Rhodes L, Rosner W, Smith RG. Sex hormone-binding globulin mediates prostate androgen receptor action via a novel signaling pathway. Endocrinology 1998;139:213-218.
10. Shiina H, Igawa M. Androgen-biosynthesis, receptor and action. Nippon Rinsho 1997; 55:2860-2864.
11. Marchetti B, Gallo F, Farinella Z, Romeo C, Morale MC. Luteinizing hormone-releasing hormone (LHRH) receptors in the neuroendocrine-immune network. Biochemical bases and implications for reproductive physiopathology. Ann N Y Acad Sci 1996; 784:209-236.
12. Moreland RB, Goldstein I, Traish A. Sildenafil, a novel inhibitor of phosphodiesterase type 5 in human corpus cavernosum smooth muscle cells. Life Sci. 1998; 62:PL.
13. Westphal RS, Anderson KA, Means AR, Wadzinski BE. A signaling complex of Ca2+-calmodulin-dependent protein kinase IV and protein phosphatase 2ª. Science 1998; 280:1258-1261.
14. Hardingham GE, Cruzalegui FH, Chawla S, Bading H. Mechanisms controlling gene expression by nuclear calcium signals. Cell Calcium 1998; 23:131-134.
15. Agell N, Aligue R, Alemany V y col. New nuclear function for calmodulin. Cell Calcium 1998; 23:115-121.
16. Waters WW, Chen PL, McArthur NH, Moreno PA, Harms PG. Calcium/calmodulin-dependent protein kinase II involvement in release of gonadotropin-releasing hormone. Neuroendocrinology 1998; 67:145-152.
17. Thurmond DC, Baillie RA, Goodridge AG. Regulation of the action of steroid/thyroid hormone receptors by medium-chain fatty acids. J Biol Chem 1998; 273:15373-15381
18. Lazar MA. Thyroid hormone receptors: Multiple forms, multiple possibilities. Endocrine Rev 1993; 14:184-193.
19. Aoki MP, Maldonado CA, Aoki A. Apoptotic and non-apoptotic cell death in hormone-dependent glands. Cell Tissue Res 1998; 291:571-574.
20. Denmeade SR, Isaacs JT. Androgen regulation of programmed (apoptotic) death in the prostate. In: Current advances in Andrology. Waites GH, Fijck J, Baker GWH (ed). Bologna:Monduzzi Editore. 1997; 143-150.
21. Kullmann M, Schneikert J, Moll J, Heck S, Zeiner M, Gehring U, Cato AC. RAP46 ia a negative regulator of glucocorticoid receptor action and hormone-induced apoptosis. J Biol Chem 1998; 273:14620-14625.
22. Mayo JC, Sainz RM, Uria H, Antolin I, Esteban MM, Rodriguez C. Melatonin prevents apoptosis induced by 6-hydroxydopamine in neuronal cells: implications for Parkinson’disease. J Pineal Res 1998; 24:179-192.
23. Shi Y, Wang R, Sharma A, Gao C, Collins M, Penn L, Mills GB. Dissociation of cytokine signals for proliferation and apoptosis. J Immunol 1997; 159:5318-5328.
24. García-Segura LM, Cardona-Gomez P, Naftolin F, Chowen JA. Estradiol upregulates Bcl-2 expression in adult brain neurons. Neuroreport 1998; 9:593-597.