STRATIGRAPHY AND AGE OF THE
MALAWI KAROO
The first published
stratigraphic section of the Chiweta Beds was that of
Dixey (1926). Total
thickness of the Karoo was measured to be 1,219 m with the upper 305 m
comprising the Chiweta Beds. Later authors have applied a K1-K7 stratigraphic
terminology devised for Tanzanian Karoo deposits, with K6 and K7 equivalent to
the Chiweta Beds, measured by
Cooper and Habgood (1959) to have a thickness of
244 m. Most bones were said to come from the middle of the unit.
Gay and
Cruickshank (1999) summarized the Tanzanian K6 and K7 units in their table 1 as Alibashian, Late Permian, Dicynodon Zone, and Anisian, middle Triassic,
Cynognathus Zone, respectively. As shown below, these designations are
too young as they are applied to the Chiweta Beds.
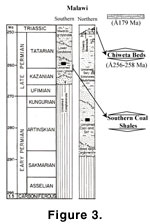 The Karoo Supergroup of Malawi
(Figure 3) exhibits no glacial or marine sediments comparable to the Dwyka Group
of South Africa. Coal-bearing strata low in the Karoo section in northern Malawi
were correlated with the South African Ecca Group by
Cairncross (2001). In
southern Malawi, Cairncross (2001) placed the coal shales in the early Tatarian
(Late Permian). However, the age of the southern Malawi coal shales is based
notably on the presence of the plant species Gangamopteris obovata
and Noeggerathiopsis hislopi, which if correct means these beds
may well be Early Permian (Habgood 1963). Given the date of the overlying Chikwawa basalt (see below) and the possible ages of plants in the coal shales,
the age range of the southern Malawi Karoo could extend from Early Permian to
Jurassic, a longer range than that presented by
Cairncross (2001, figure 2;
Figure 3).
The Karoo Supergroup of Malawi
(Figure 3) exhibits no glacial or marine sediments comparable to the Dwyka Group
of South Africa. Coal-bearing strata low in the Karoo section in northern Malawi
were correlated with the South African Ecca Group by
Cairncross (2001). In
southern Malawi, Cairncross (2001) placed the coal shales in the early Tatarian
(Late Permian). However, the age of the southern Malawi coal shales is based
notably on the presence of the plant species Gangamopteris obovata
and Noeggerathiopsis hislopi, which if correct means these beds
may well be Early Permian (Habgood 1963). Given the date of the overlying Chikwawa basalt (see below) and the possible ages of plants in the coal shales,
the age range of the southern Malawi Karoo could extend from Early Permian to
Jurassic, a longer range than that presented by
Cairncross (2001, figure 2;
Figure 3).
The
Age of the Chikwawa Basalt
The distribution of the
Drakensberg Volcanics and related rocks that cap the Karoo Supergroup defines
the Karoo Large Igneous Province (LIP). Karoo LIP outcrops extend on the east
from the Drakensberg Mountains of South Africa and Lesotho, through the Lebombo
Mountains of Swaziland and Mozambique. They include the Etjo basalt of Namibia
and the Batoka basalts of
Zimbabwe. By proximity to the Lebombo volcanics and the Batoka basalts, the
older set of extrusive rocks of the Lupata Gorge in Mozambique (Dixey and Smith
1929), and the Chikwawa basalt of southern Malawi appear to be the northern
outcrop limit of the Karoo LIP.
The thickness of
the igneous rocks capping the Karoo Chikwawa Group of southern Malawi is
estimated by Habgood (1963) to be 1067 m, but exposures are poor, faulting
complicates the relationship of outcrops, and the age is poorly constrained.
Nevertheless, as in Karoo extrusives in other areas, weathered surfaces between
flows are rare, suggesting a rapid series of eruptions (Habgood 1963).
Based on geochronology and paleomagnetism of the
Batoka basalts in northern Zimbabwe,
Jones et al. (2001) determined that the
duration of emplacement of the Karoo LIP was
≈5
Myr, ranging between approximately 184 to 179 Ma.
 We report here the
results of laser incremental-heating 40Ar/39Ar
age determinations of a
Chikwawa basalt flow (sample LJ-4) and a cross-cutting basaltic dike (LJ-3) from
samples collected in 1990. These rocks are nearly identical, fine-grained
intergranular plagioclase and pyroxene basalts showing minor alteration of
mineral phases in thin section. The flow overlies Karoo Supergroup sediments on
the south side of the Nyakamba River in a roadcut for Highway M8 (Figure 1,
Figure 4)
near the town of Ngabu (1627¥20¥¥S, 3443¥45¥¥E).
We report here the
results of laser incremental-heating 40Ar/39Ar
age determinations of a
Chikwawa basalt flow (sample LJ-4) and a cross-cutting basaltic dike (LJ-3) from
samples collected in 1990. These rocks are nearly identical, fine-grained
intergranular plagioclase and pyroxene basalts showing minor alteration of
mineral phases in thin section. The flow overlies Karoo Supergroup sediments on
the south side of the Nyakamba River in a roadcut for Highway M8 (Figure 1,
Figure 4)
near the town of Ngabu (1627¥20¥¥S, 3443¥45¥¥E).
 Two aliquots of
crushed (0.15 to 0.25 mm) whole rock were analyzed from each of the two samples
(Figure 5, see
Table 1,
Deino et al. 1990, and
Sharp et al. 1996,
for details of the analytical procedure). Reproducibility of fine details of
release patterns in age and geochemical parameters (%40Ar*, Ca/K),
and in the derived plateau and integrated ages between aliquots of the same
sample, is noteworthy. All spectra released initial fractions (<10% of the
cumulative 39Ar release) charged with atmospheric argon and younger
than the subsequent plateau, probably reflecting a limited degree of alteration.
Nevertheless, broad plateaus occur within the central portion of the release
sequence. These plateaus occupy the zone where the Ca/K ratio of the released
gasses is less than about 4. In the final 20-30% of the 39Ar release,
Ca/K ratios rise sharply and apparent ages fall. This spectral form,
characterized by a stair-stepping downward pattern in both the beginning and end
of the incremental-heating sequence, is consistent in all four experiments. It
can be interpreted as representing recoil implantation of 39Ar
released from fine-grained alteration phases into refractory calcic phases. The
fine-grained, 39Ar depleted alteration products release their argon
early and exhibit artificially old ages and high atmospheric content, while the
39Ar enriched calcic phases retain their argon to the last,
exhibiting young ages and high Ca/K ratios. Despite this redistribution of
39Ar locally, older apparent ages in the early release steps are balanced
against young ages in the final steps, so that integrated ages are in all cases
congruent with the plateau results.
Two aliquots of
crushed (0.15 to 0.25 mm) whole rock were analyzed from each of the two samples
(Figure 5, see
Table 1,
Deino et al. 1990, and
Sharp et al. 1996,
for details of the analytical procedure). Reproducibility of fine details of
release patterns in age and geochemical parameters (%40Ar*, Ca/K),
and in the derived plateau and integrated ages between aliquots of the same
sample, is noteworthy. All spectra released initial fractions (<10% of the
cumulative 39Ar release) charged with atmospheric argon and younger
than the subsequent plateau, probably reflecting a limited degree of alteration.
Nevertheless, broad plateaus occur within the central portion of the release
sequence. These plateaus occupy the zone where the Ca/K ratio of the released
gasses is less than about 4. In the final 20-30% of the 39Ar release,
Ca/K ratios rise sharply and apparent ages fall. This spectral form,
characterized by a stair-stepping downward pattern in both the beginning and end
of the incremental-heating sequence, is consistent in all four experiments. It
can be interpreted as representing recoil implantation of 39Ar
released from fine-grained alteration phases into refractory calcic phases. The
fine-grained, 39Ar depleted alteration products release their argon
early and exhibit artificially old ages and high atmospheric content, while the
39Ar enriched calcic phases retain their argon to the last,
exhibiting young ages and high Ca/K ratios. Despite this redistribution of
39Ar locally, older apparent ages in the early release steps are balanced
against young ages in the final steps, so that integrated ages are in all cases
congruent with the plateau results.
We take the
weighted means of the plateau ages of the replicates as the reference age, and
obtain 179.4 ± 0.8 for LJ-3 and 168.6 ± 0.8 for LJ-4. These determinations are
clearly distinct and present the apparently inverted age relationship of an
older age for the crosscutting basalt than the lava flow it lies within. Either
alteration has affected the argon systematics more than is apparent from the
spectra and the lava flow is relatively too young, or the dike contains a
component of excess argon and is relatively too old.
In either case,
these ages are relatively young, but broadly compatible with the emplacement
history of the Karoo LIP (184 to 179 Ma,
Duncan et al. 1997;
Jones et al. 2001).
Considering the geographic proximity of the Chikwawa basalt relative to
recognized Karoo LIP rocks, the stratigraphic position of the Chikwawa basalt
atop Karoo Supergroup sediments, and the age of the lava flow and dike at Ngabu,
the Chikwawa basalt is taken to be the northern outcrop extent of the Karoo LIP.
The age of the Chikwawa basalt is Jurassic, and in the absence of a major
unconformity beneath this flow, the Karoo strata of southern Malawi are much
younger than the Triassic age assigned by
Cairncross (2001, figure 2;
Figure 3).
Northern
Malawi
The Chiweta Beds of northern
Malawi are fluvial in origin. Bones within overbank deposits are often found
encased in pedogenic carbonate, similar to the condition of those from the
Beaufort Group of South Africa (Smith 1990,
1993). The most detailed stratigraphic work conducted on the Karoo Supergroup of the Mount Chombe-Chiweta
area is that by
Yemane et al. (1989),
Yemane and Kelts (1990), and
Yemane
(1993). Approximately 300 m of section were measured, all underlying the therapsid-bearing Chiweta Beds, which were excluded from their studies.
Yemane et al. (1989) interpreted the environment of deposition to be lacustrine, and
while these lacustrine sediments do not preserve bone, they contain pollen
(Yemane 1994). Of 20 identified pollen taxa, 15 are unknown after the
Permian-Triassic boundary. Eleven of those taxa have records prior to the Late
Permian, and four range throughout the Late Permian. Three pollen taxa (Taeniaesporites
noviaulensis, Lycopodiacidites pelagius, and
Rimaesporites aquilonalis) range only from after the beginning of the
Tatarian (late Capitanian or younger fide
Gradstein and Ogg 2004) and
into the Triassic. Therefore, an independent maximum age limit for the overlying Chiweta Beds of <263 Ma (base of the Tatarian,
timescale of
Gradstein and Ogg 2004) is set from palynological evidence.
Age of the Chiweta Beds Based
on Therapsid Biostratigraphy.
Dixey (1926) recognized two main bone
producing horizons in the Chiweta Beds, although he acknowledged that bone could
be found throughout. The main bone producing horizons correspond to his units 5
and 7. Unit 5 is the Lower Bone Bed (or B1), and unit 7 is the Upper Bone Bed
(B2) of Haughton (1926). Our collections were made from the Lower Bone Bed (B1,
seen in the foreground in Figure 2) with the possible exception of an
articulated but unprepared and unidentified skeleton lacking the skull, which is
likely from the Upper Bone Bed (B2).
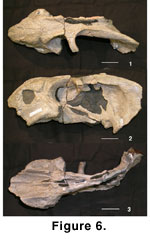 The upper age limit of the
Chiweta Beds can be refined by comparison of the B1 therapsid assemblage with
that of the Beaufort Group, the vertebrate biostratigraphy of which is reviewed
in Rubidge (1995). Of particular importance is the genus Oudenodon, which
we show to occur at Chiweta (Figure 6,
Figure 7). King diagnosed the Tribe Oudenodontini (which contained only Oudenodon) as:
The upper age limit of the
Chiweta Beds can be refined by comparison of the B1 therapsid assemblage with
that of the Beaufort Group, the vertebrate biostratigraphy of which is reviewed
in Rubidge (1995). Of particular importance is the genus Oudenodon, which
we show to occur at Chiweta (Figure 6,
Figure 7). King diagnosed the Tribe Oudenodontini (which contained only Oudenodon) as:
Medium-sized to large
dicynodonts (skull length ranging from 100 mm to over 300 mm). Teeth lacking in
both upper and lower jaws. Postorbitals well separated on skull roof by
parietals. Septomaxilla recessed within external naris, lachrymal in some
species extends forward above maxilla to posterior margin of naris. Nasal forms
boss over naris. Maxilla carries weak caniniform process, with sharp edged
posterior crest. Palatal portion of palatine divided into inflated posterior
area, and a smooth anterior part that meets the premaxilla. Vomers form short
septum in anterior part of interpterygoidal fossa. Ectopterygoid large with
palatal exposure, pterygoid does not contact maxilla… (King 1988, p. 85).
 The characters observed in the
dicynodont skulls designated Malawi Department of Antiquities (Mal) 108 and 129
(Figure 6,
Figure 7) from the Chiweta Beds indicate that both belong to the genus
Oudenodon. Mal 108 is lacking its posterior portion. Mal 129 is
postdepositionally dorsoventrally compressed. Measurements (Table 2) demonstrate
that the two specimens are similar in size and represent a single species, but
Mal 129 appears more robust.
The characters observed in the
dicynodont skulls designated Malawi Department of Antiquities (Mal) 108 and 129
(Figure 6,
Figure 7) from the Chiweta Beds indicate that both belong to the genus
Oudenodon. Mal 108 is lacking its posterior portion. Mal 129 is
postdepositionally dorsoventrally compressed. Measurements (Table 2) demonstrate
that the two specimens are similar in size and represent a single species, but
Mal 129 appears more robust.
Oudenodon groups with
Tropidostoma, Rhachiocephalus, and their most recent common ancestor
within a clade (Cryptodontinae) within the Dicynodontidae (sensu
King 1988,
1990;
Angielczyk 2001; compare
Angielczyk and Kurkin 2003a,
2003b). Members of this clade are diagnosed by having a postcaniniform crest. A less inclusive clade
within the Cryptodontinae, diagnosed by parietals exposed in a depression
between the postorbitals and a long interpterygoid vacuity, includes
Tropidostoma and Oudenodon, but not Rhachiocephalus
(Angielczyk 2001). Of these taxa, teeth are present in Tropidostoma but
lacking in Oudenodon.
Haughton (1926) reported two
species of Dicynodon distinguished by size in the material sent him by
Dixey from Chiweta. Both are tuskless. The illustrations in
Haughton (1926) of
the smaller species, designated by him Dicynodon sp. A, show the
parietals exposed between the postorbitals, indicating that it should be
referred to Oudenodon. The larger species was represented by a
fragmentary skull, a humerus, and parts of other postcranial bones, which
Haughton designated Dicynodon sp. cf. D. grandis. The referral of
those specimens to D. grandis, a species named by Haughton (1917), was
based on size alone. Keyser
(1975), in his review of tuskless anomodonts,
referred D. grandis to the genus Oudenodon, consolidating
the genus and synonymizing a large number of species into three: O. baini,
the genotype, most abundant, and widespread species; O. grandis, a rare
but large species whose only diagnostic character other than size is having the
parietal foramen lie in a depression on the dorsal surface of the skull; and
O. luangwaensis, known from Zambia and defined on the basis of “…the great
width of the zygomatic arches, giving the skulls their very characteristic heart
shape” (Keyser 1975, p. 57).
Measurements of Mal 108 and Mal
129 (Table 2) fall within the range of Oudenodon baini presented by
Keyser (1975, p. 39-40) indicating that those specimens do not pertain to O.
grandis. In addition, a low boss lies near the parietal foramen of Mal 108,
which if a reliable character, also indicates that Mal 108 is not O. grandis.
Mal 129 lacks the great width and therefore the “very characteristic heart
shape” of the skull of O. luangwaensis, even though this character
in Mal 129 is accentuated by dorsoventral flattening. Haughton’s Dicynodon
sp. A is smaller than either Mal 108 or Mal 129, but still within the range of
O. baini. We therefore identify Mal 108, 129, and Dicynodon sp. A
of Haughton (1926) as Oudenodon baini.
Haughton (1926) also reported
teeth and jaw fragments of Endothiodon cf. E. bathystoma. He
questioned the level from which they were obtained but considered them most
likely to come from the Lower Bone Bed (B1), and therefore less likely to have
come from the Upper Bone Bed (B2). We have not recorded Endothiodon in
the collections we made from B1. Regardless, Endothiodon is a distinctive
genus (Cox 1964) and its occurrence in the Chiweta Beds has biostratigraphic
significance. Haughton (1926, p. 82), in evaluating the age of the Chiweta
fauna, states, “…had [the assemblage] been found in South Africa, [it] would
unhesitatingly have been assigned to the base of the Cistecephalus zone
or top of the Endothiodon zone of the Lower Beaufort Beds.”
Numerous revisions to the
biostratigraphy of the Beaufort Group have been made since Haughton’s work, but
his observations are on the mark. In the latest comprehensive revision,
Endothiodon is a component of the upper Pristerognathus Assemblage
Zone (Smith and Keyser 1995a), the Tropidostoma Assemblage Zone (Smith
and Keyser 1995b), and at least the lower portion of the Cistecephalus
Assemblage Zone (Smith and Keyser, 1995c). Oudenodon is found throughout
the Cistecephalus and the overlying Dicynodon assemblage zones
(Kitching 1995), becoming extinct near the Permian-Triassic boundary.
Gorgonops is known from the Tropidostoma and most of the
Cistecephalus Assemblage Zones (Smith and Keyser 1995b). Thus, the
overlapping ranges of Endothiodon and Oudenodon, consistent with
the presence of Gorgonops? dixeyi and Aelurognathus, argue
for a lower Cistecephalus Assemblage Zone placement for the Chiweta Beds.
The youngest likely age falls just below the midpoint of the Tatarian Stage
(Hancox and Rubidge 2001), or following
Gradstein and Ogg (2004), about at the
middle of the Wuchiapingian Stage, or approximately 256 to 258 Ma. We take that
range as a best estimate for the age of the fauna.
In conjunction with his
phylogenetic analyses of dicynodonts,
Angielczyk (2001,
2002);
Angielczk and
Kurkin (2003a,
2003b) fitted preferred cladograms to the stratigraphic record of
the taxa, thereby demonstrating the extent of ghost lineages associated with
most parsimonious trees. The most significant implication for this study affects
the overlap between Endothiodon and Oudenodon, a co-occurrence
that limits the correlation of the Chiweta Beds with the Cistecephalus
Assemblage Zone, but eliminates a correlation with the uppermost portion of that
same zone. The downward ghost extension of Oudenodon, it might be argued,
could affect the correlation of the Chiweta Beds with the Karoo Basin of South
Africa by allowing a correlation with the underlying Tropidostoma
Assemblage Zone. However, the biostratigraphic ranges of the robust suite of
pollen taxa (Yemane 1994) limit the maximum age of the therapsid assemblage and
mitigate concern over downward extension of the Oudenodon range.
