INTRODUCTION
Interpretations of the functional
morphology of fossil cephalopods have tended to concentrate on
those that resemble extant cephalopods (e.g., belemnites compared
to squid, Monks et al. 1996; regularly coiled ammonites to Nautilus,
Chamberlain 1976, 1980).
Fossil cephalopods that do not have such clear analogues, such as
the Lower Cretaceous heteromorph ammonites, are more
problematical. Moreover, it is unclear how far Nautilus is
a valid analogue for ammonites.
Nautilus has paired
retractor muscles that can pull the head and arms into the shell
when the animal is threatened. The aperture is also sealed by a
tough hood, functionally similar to the gastropod operculum (Wells et al. 1992). However, the Nautilus body chamber
is short, approximately one third of a whorl in length, and the
animal cannot withdraw far. In contrast, the living chambers of
ammonites are very long and often narrow. Moreover, ammonites
lacked a hood or operculum. [Note: the aptychi are now generally
accepted to have been part of the jaws (Lehmann 1981)]. Consequently, the assumption that the
soft parts of the ammonite filled the entire living chamber of
the shell is not neccessarily correct, and it is possible that
without a way of sealing the aperture, ammonites may have been
capable of withdrawing further into the shell than has heretofore
been acknowledged.
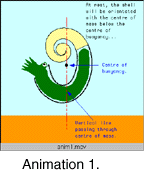 Body
position in heteromorph ammonites is important because the
association of the soft body (e.g., head, arms, viscera) with the
shell is a key part of understanding how these animals lived and
functioned. The discussion by Trueman (1941) on this topic is widely quoted (e.g., in
the Treatise on Invertebrate Paleontology, Arkell et al. 1957). Trueman assumed that the entire living
chamber was filled, and used this to deduce the
centres of mass and buoyancy of the ammonite. Ammonite functional
interpretations based on this model are characterised by a single
stable orientation, with the centre of mass vertically below the
centre of buoyancy. For heteromorph ammonites this model
typically predicts upturned apertures (Anim.
1). Many workers have cited
these upturned apertures as evidence that heteromorphs were
pelagic plankton feeders (e.g., Klinger 1980, Westermann 1996).
Body
position in heteromorph ammonites is important because the
association of the soft body (e.g., head, arms, viscera) with the
shell is a key part of understanding how these animals lived and
functioned. The discussion by Trueman (1941) on this topic is widely quoted (e.g., in
the Treatise on Invertebrate Paleontology, Arkell et al. 1957). Trueman assumed that the entire living
chamber was filled, and used this to deduce the
centres of mass and buoyancy of the ammonite. Ammonite functional
interpretations based on this model are characterised by a single
stable orientation, with the centre of mass vertically below the
centre of buoyancy. For heteromorph ammonites this model
typically predicts upturned apertures (Anim.
1). Many workers have cited
these upturned apertures as evidence that heteromorphs were
pelagic plankton feeders (e.g., Klinger 1980, Westermann 1996).
 More
recently, Kakabadzé and
Sharikadzé (1993) proposed
that the heteromorph shell had two stable orientations, one with
the aperture pointing upwards (as in previous interpretations)
and a second with the aperture tilted towards the sea floor. This
alternative orientation would make it possible for heteromorph
ammonites to feed off the sea floor. Kakabadzé and
Sharikadzé (1993) also
proposed that these ammonites might have been able to adjust
their bouyancy by shifting cameral fluid within the shell. This
would change the position of the centre of mass, and consequently
the orientation of the shell (Anim. 2).
More
recently, Kakabadzé and
Sharikadzé (1993) proposed
that the heteromorph shell had two stable orientations, one with
the aperture pointing upwards (as in previous interpretations)
and a second with the aperture tilted towards the sea floor. This
alternative orientation would make it possible for heteromorph
ammonites to feed off the sea floor. Kakabadzé and
Sharikadzé (1993) also
proposed that these ammonites might have been able to adjust
their bouyancy by shifting cameral fluid within the shell. This
would change the position of the centre of mass, and consequently
the orientation of the shell (Anim. 2).
The idea that ammonites may have
had more than one stable orientation has not been widely
discussed by ammonite workers, but if true could force a
re-appraisal of ammonite paleoecology. The Kakabadzé and
Sharikadzé (1993) model
relies on ammonites being able to rapidly change the distribution
or amount of fluid within the shell. As far as is known, movement
of fluid within the shell of ectocochleates is slow. In Nautilus,
changes in the amount of fluid within the chambers is too slow
even to assist diurnal migration by adjusting overall buoyancy
(Chamberlain 1991). 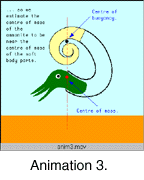 Cuttlefish
have a highly modified and characteristic shell, divided into
numerous small chambers, subdivided by conchiolin walls, and
exhibiting a broad siphuncular region rather than a narrow
siphuncle (Denton & Gilpin-Brown
1961a). This appears to
allow the cuttlefish to make changes to the buoyancy of the shell
more quickly than Nautilus (Denton & Gilpin-Brown
1961b). Even so, buoyancy
regulation in cuttlefish seems not to be used as the primary
device for rising or sinking in the water column, but rather for
acquiring neutral buoyancy once a preferred depth has been
reached (Denton 1973). Ammonite shells are quite unlike the
shells of cuttlefish and so it is more likely that Nautilus
is an appropriate analogue in this respect. As a result, the
problem remains that while changes in orientation may have been
possible, no entirely convincing mechanism has been proposed for
how this might have been acomplished. We propose a third possible
anatomy: That the heteromorph ammonite animal was a relatively
small, mobile creature that was able to move within its floating
shell rather like a modern gastropod. As with the previous model
this allows significant changes in orientation, due in this case
to changes in the distribution of the mass as the body moves
within the shell (Anim. 3).
Cuttlefish
have a highly modified and characteristic shell, divided into
numerous small chambers, subdivided by conchiolin walls, and
exhibiting a broad siphuncular region rather than a narrow
siphuncle (Denton & Gilpin-Brown
1961a). This appears to
allow the cuttlefish to make changes to the buoyancy of the shell
more quickly than Nautilus (Denton & Gilpin-Brown
1961b). Even so, buoyancy
regulation in cuttlefish seems not to be used as the primary
device for rising or sinking in the water column, but rather for
acquiring neutral buoyancy once a preferred depth has been
reached (Denton 1973). Ammonite shells are quite unlike the
shells of cuttlefish and so it is more likely that Nautilus
is an appropriate analogue in this respect. As a result, the
problem remains that while changes in orientation may have been
possible, no entirely convincing mechanism has been proposed for
how this might have been acomplished. We propose a third possible
anatomy: That the heteromorph ammonite animal was a relatively
small, mobile creature that was able to move within its floating
shell rather like a modern gastropod. As with the previous model
this allows significant changes in orientation, due in this case
to changes in the distribution of the mass as the body moves
within the shell (Anim. 3).
 The
only direct evidence for soft part morphology in ammonites is
provided by the muscle scars. These are consistently located near
the rear of the living chamber, close to the final septum (Crick 1898). Such a muscular arragement is compatible
with our model of a small retractable body although clearly it
does not prove it. Heteromorph ammonite shells are characterized
by a departure from the regular planispiral shape of most other
ammonites. One of the most common themes in heteromorph shell
design is that of the planispiral phragmocone and hook-shaped
living chamber.
The
only direct evidence for soft part morphology in ammonites is
provided by the muscle scars. These are consistently located near
the rear of the living chamber, close to the final septum (Crick 1898). Such a muscular arragement is compatible
with our model of a small retractable body although clearly it
does not prove it. Heteromorph ammonite shells are characterized
by a departure from the regular planispiral shape of most other
ammonites. One of the most common themes in heteromorph shell
design is that of the planispiral phragmocone and hook-shaped
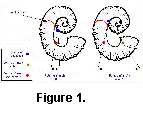
living chamber.  The
simplest form is that of the aspinocone, where the living chamber
consists of a shallow U-shaped hook held slightly away from the
planispiral phragmocone (Fig. 1). By drawing the hook deeper into two parallel
shafts, the ancyclocone is produced (Fig. 2). Finally, straightening all but the earliest
stages of the phragmocone and tightening the bends further
results in the hamiticone (Fig. 3).
The
simplest form is that of the aspinocone, where the living chamber
consists of a shallow U-shaped hook held slightly away from the
planispiral phragmocone (Fig. 1). By drawing the hook deeper into two parallel
shafts, the ancyclocone is produced (Fig. 2). Finally, straightening all but the earliest
stages of the phragmocone and tightening the bends further
results in the hamiticone (Fig. 3).