TAXONOMY
Outline classification
The following synopsis, based
chiefly on the scheme proposed by Riedel (1967b, 1971),
is restricted to extant families only. As pointed out above, it
is a provisional classification very likely to change as more
structural and evolutionary data are obtained. Taxa in bold
characters are those treated in detail in this chapter.
Kingdom PROTISTA
Haeckel, 1886
Phylum SARCODINA Hertwig and Lessser, 1876
Class ACTINOPODA Calkins, 1909
Subclass HELIOZOA Haeckel, 1886
Subclass ACANTHARIA Müller, 1858
Subclass RADIOLARIA Müller, 1858
Superorder PHAEODARIA Haeckel, 1879
Superorder POLYCYSTINA Ehrenberg, 1838, emend. Riedel,
1967
Order COLLODARIA Haeckel, 1881
Family
THALASSICOLLIDA Haeckel, 1862
Family COLLOZOIDA Haeckel, 1862
Family THALASSOSPHAERIDA Haeckel, 1862
Family SPHAEROZOIDA Haeckel, 1862
Order SPUMELLARIA
Ehrenberg, 1875
Family COLLOSPHAERIDAE
Müller, 1858, emend. Strelkov and Reshetnjak, 1971
Family ACTINOMMIDAE Haeckel, 1862, emend.
Sanfilippo and Riedel, 1980
Family COCCODISCIDAE Haeckel, 1862 emend.
Sanfilippo and Riedel, 1980
Family PHACODISCIDAE Haeckel, 1881
Family SPONGODISCIDAE Haeckel, 1862, emend. Riedel
1967
Family LITHELIIDAE Haeckel, 1862
Family PYLONIIDAE Haeckel, 1881
Family THOLONIIDAE Haeckel, 1862
Order NASSELLARIA
Ehrenberg, 1875
Family SPYRIDAE
(=Trissocyclidae, Acanthodesmiidae) Ehrenberg, 1847,
emend. Petrushevskaya, 1971
Family PLAGONIIDAE Haeckel, 1881, emend. Riedel,
1967
Family THEOPERIDAE Haeckel, 1881, emend. Riedel,
1967
Family CARPOCANIIDAE Haeckel, 1881, emend. Riedel,
1967
Family PTEROCORYTHIDAE Haeckel, 1881, emend.
Riedel, 1967
Family ARTOSTROBIIDAE Riedel, 1967, emend.
Foreman, 1973
Family CANNOBOTRYIDAE Haeckel, 1881, emend.
Riedel, 1967
Order and family-level diagnoses
Order Collodaria. Solitary
or colonial polycystines without a siliceous skeleton, or
provided with simple or branched spicules scattered in the
calymma. Due to their fragility, members of this group preserve
poorly in net plankton samples, and either do not preserve at all
or are represented only by their spicules in sedimentary
materials. Partly because of these limitations, information on
their classification and distribution is extremely scarce, and no
further details are given herein. Detailed reviews of the
colonial radiolarians, including several Collodaria, were
produced by Hollande and Enjumet (1953), Strelkov and Reshetnjak
(1971), and Swanberg (1979). Most of these species have tropical
distribution ranges in the three major oceans. In the south
Atlantic they are probably restricted to waters associated with
the equatorial current system, the Tropics/Subtropics, and the
oligotrophic Central Gyre (Figure 11). According to Haeckel (1887), this group comprises four families:
Family Thalassicollida:
solitary cells, no skeletal elements; genera Actissa
(Figure 15.1), Thalassocampe (Figure 15.2), Thalassopila,
Thalassicolla, and Thalassophysa;
Genus Collozoida:
colonial, no skeletal elements; genus Collozoum
(Figure 15.6, 15.7, 15.8, 15.11);
Genus Thalassosphaerida:
solitary, with siliceous spicules scattered in the calymma;
genera Thalassosphaera, Thalassoxanthium
(Figure 15.3), Physematium, Thalassoplancta, Lampoxanthium
(Figure 15.5);
Genus Sphaerozoida:
colonial, with siliceous spicules scattered in the calymma;
genera Belonozoum, Sphaeorozoum (Figure 15.10),
Raphidozoum (Figure 15.12).
Order Spumellaria. Solitary
or colonial radiolarians with a well-developed shell of radial
symmetry or one derived from the above. Variations in the type of
symmetry include spiral shells (e.g., Figure 15.87), asymmetric,
discoidal or lenticular (biconvex) (Figure 2H, 2I; 15.60, 15.61,
15.62, 15.63, 15.64, 15.65), triaxonic (Figure 2Q, 2R),
quadrangular (Figure 15.66, 15.67), etc. In many cases two axes
of symmetry can be clearly differentiated (Figure 2J) but, as
opposed to the Nassellaria, the larger axis is homoaxonic. The
central capsule (organic) of these cells has many small pores.
Family Collosphaeridae.
Colonial polycystines, each individual has a single,
thin-walled, spherical or subspherical latticed shell. The
Collosphaeridae is the only group of colonial polycystines
with complete latticed shells. Colonies consist of a
gelatinous mass (which obviously disappears in the
sedimentary record, as well as in many net-plankton samples
where it breaks down) in which hundreds to thousands of
shells are immersed (Figure 1C, 1D, 1E).  The shape of the colony is not
species-specific; it may be spherical, ellipsoidal,
cylindrical, ribbon-shaped, etc., measuring up to several
centimeters in length and a few millimeters in diameter. The
siliceous shells are always represented by a single
perforated sphere (internal spheres are never present), with
or without centrifugal (external) or centripetal (internal)
tubular projections and/or spines. Spines (when present) are
conical (circular in cross-section). As with most other
polycystines, specific assignments are based almost
exclusively on the skeleton; however, studies of entire
colonies, which allow investigating the intraspecific
morphologic variability of the collosphaerids, indicate that
quite dissimilar shell morphotypes can coexist within the
same colony, thus stressing the assumption that at least some
of the specific divisions based on the siliceous sphere alone
are spurious (e.g., Kleijne 1987; Petrushevskaya and
Swanberg 1990).
The shape of the colony is not
species-specific; it may be spherical, ellipsoidal,
cylindrical, ribbon-shaped, etc., measuring up to several
centimeters in length and a few millimeters in diameter. The
siliceous shells are always represented by a single
perforated sphere (internal spheres are never present), with
or without centrifugal (external) or centripetal (internal)
tubular projections and/or spines. Spines (when present) are
conical (circular in cross-section). As with most other
polycystines, specific assignments are based almost
exclusively on the skeleton; however, studies of entire
colonies, which allow investigating the intraspecific
morphologic variability of the collosphaerids, indicate that
quite dissimilar shell morphotypes can coexist within the
same colony, thus stressing the assumption that at least some
of the specific divisions based on the siliceous sphere alone
are spurious (e.g., Kleijne 1987; Petrushevskaya and
Swanberg 1990).
Family Actinommidae.
Solitary species with latticed or spongy spherical,
subspherical, or ovoid shells (not lenticular); with or
without medullary shells. Surface of shell is often covered
with spines, but not tubes. All actinommids posses either
single or multiple, concentric spherical or ovoid shells.
When several shells are present they are connected to each
other by radial beams which pierce the cell. An enormous
variety of forms was described in this family whose
identification has traditionally been based on Haeckel's (1887) system. Haeckel based the
classification of the actinommids (=suborder Sphaeroidea,
exclusive of the Collosphaerida) on the following characters
(in decreasing order of importance; see Figure 13): 1. Number
of primary radial spines; 2. Number of concentric spheres; 3.
Position of concentric spheres (intra- or extracapsular),
type and relative size of spines, presence of by-spines, type
of medullary shell, etc.). However, the number of primary
spines varies intraspecifically, whereas the number of main
concentric spheres, which within some bounds might indeed be
species-specific (Riedel and Sanfilippo
1986), can only be used
in the case of fully-grown individuals. It is quite obvious
that, based on this trait, Haeckel (as well as many other
authors) assigned new names to growth stages still missing
the outermost sphere(s) (see Figs. 13 and 14). Furthermore,
while growth of an actinommid as far as we know proceeds from
the center toward the periphery (Figure 14, upper panel),
dissolution works in the opposite direction, innermost, more
delicate shells usually disappearing before the more robust
cortical ones. Thus, materials from the sediments offer yet
another suite of "new species", this time missing
the medullary (rather than the cortical) shells.
Family Coccodiscidae
(Figure 2H, 2J). Latticed discoidal or lenticular shell
enclosing a single or double medullary shell, and surrounded
by an equatorial zone of spongy or concentrically-chambered
structures (Figure 2H), or forms with an ellipsoidal cortical
shell equatorially constricted enclosing a single or double
medullary shell (Figure 2J). The formerly actinommid
subfamily Artiscinae was transferred to the Coccodiscidae by Sanfilippo and Riedel
(1980) due to its
phylogenetic affinities with extinct coccodiscids.
Family Phacodiscidae.
Lenticular, biconvex, latticed cortical shell, not surrounded
by spongy or chambered structures, within which a small,
spherical single or double medullary shell is enclosed. The
margin (but less commonly the surfaces) of the cortical shell
may bear radial spines.
Family Spongodiscidae.
Discoidal or cylindrical, spongy or finely chambered
skeleton, with or without surficial pore-plate, often with
radiating arms or marginal spines. The members of this family
are characterized by possessing skeletons which are partly or
entirely spongy in appearance. However, as opposed to the
Actinommidae, which can also have spongy skeletons, the
Spongodiscidae are not spherical. Their overall shape can be
lenticular (biconvex discs, Figure 2I), cylindrical (Figure
15.74), quadrangular or subquadrangular in outline (Figure
15.67), or Y-shaped (Figure 2Q, 2R). With the exception of
the cylinders, all others are depressed or flattened (rather
than circular in cross-section, Figure 2I). Lenticular,
quadrangular, and Y-shaped forms may be entirely composed of
a spongy mass with no discernible structure (in which case
the central part of the skeleton is often thicker and/or
denser, and therefore appears darker in the light microscope;
Figure 2O, 15.64), or may posses a small central chamber
surrounded by concentric or spiral, continuous or interrupted
bands (Figure 2R). The surface of some forms may be partly or
totally covered with a very thin, porous sieve-plate, which
in lenticular forms may extend beyond the central spongy mass
forming a delicate equatorial girdle around the periphery of
the shell (Figure 2P) (these morphotypes were formerly
included in the family Porodiscidae).
Family Litheliidae. The
lattice of the ellipsoidal, spherical or lenticular shell is
totally or partially arranged along a bilaterally symmetrical
spiral. Although very abundant, due to their complicated
architecture the litheliids are poorly known, for which
reason the morphotypes defined may include several different
forms.
Family Pyloniidae. The
major part of the shell is composed of a series of
successively larger elliptical latticed girdles in three
mutually perpendicular planes, with the major diameter of
each girdle being the minor diameter of the next larger one
(Figure 2L, 2M). The center is occupied by a small
ellipsoidal structure - the microsphere (see Dumitrica 1989).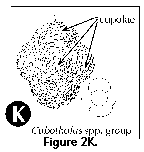
Family Tholoniidae.
Completely latticed shell, without larger openings, and with
constrictions that define several (typically 6) dome-shaped
protuberances (Figure 2K).
Order Nassellaria. Solitary
polycystines with a siliceous heteropolar shell, which can be
represented by several fused spicules only, by a D-shaped ring
and associated spines, or by more elaborate, mono- or
multilocular latticed skeletons. With the exception of a few
forms lacking a well developed skeleton (Figure
15.101, 15.119), the symmetry of this group is
characterized by the fact that the two extremes of the major axis
define two morphologically different poles of the shell. One of
these, conventionally accepted as the top or anterior end, is
where the cephalis is located. A widely recognized, albeit seldom
utilized, feature of primary importance for the classification of
the Nassellaria is the internal skeleton. The internal skeleton
consists of a complex set of spines and connecting bars enclosed
in the cephalis (Figure 3A, 3B, 3C), which allow comparison of
homologous structures in forms differing widely in their external
morphology. Unfortunately, analysis of these features requires
dedicated efforts at understanding the complex spatial
relationships involved. Furthermore, observation of this internal
skeleton is only feasible with well preserved individuals
oriented in the right position, which is seldom the case in
specimens mounted in permanent slides. In addition to the small
scattered perforations typical of the Spumellaria, the central
capsule of the Nassellaria is usually provided with a single
larger pore.
Family Spyridae
(=Trissocyclidae). The skeleton is represented by a
well-developed D-shaped sagittal ring (median bar and
anastomosed vertical and apical spines), either free (Figure
15.101) or embedded into the latticed cephalic wall, in which
case the cephalis is usually bilaterally lobed (Figure 15.93,
15.94, 15.100). Sometimes with thorax, abdomen always absent.
The typical heteropolar nassellarian symmetry is often
inconspicuous in the Spyridae.
Family Plagoniidae.
Skeletons restricted to a simple tri- or tetraxonic
nassellarian spicule (Figure 15.119), or a well developed
system of main spines enclosed within a fully formed cephalis
(Figure 3A, 3B). The degree of development of the cephalis
may vary from a few anastomosed bars (Figure 15.120, 15.124)
to a well developed, latticed or latticed/spongy chamber.
Usually without postcephalic segments. In addition to several
fairly well-defined species, the Plagoniidae comprise many
probably related forms of obscure taxonomic status usually
cited under various generic names (see below). The
classification of these forms needs detailed ad hoc studies,
for which reason many of them are provisionally lumped under
the designation Plagoniidae group in the present
chapter.
Family Theoperidae.
Cephalis spherical or subspherical, relatively small, often
poreless or sparsely perforate. It usually bears an apical
horn. Internal spicule small and inconspicuous. With one or
more, sometimes up to over 10, usually well-developed
postcephalic segments. Generally, cap- or helmet-shaped, or
conical in overall outline.
Family Carpocaniidae.
The small, rudimentary cephalis is usually totally immersed
in the large and well-developed thorax (Figure 3E). Abdomen
absent or rudimentary.
Family Pterocorythidae.
Cephalis large, divided into three lobes by two lateral
furrows directed obliquely and downward from the apical spine
to the base of the cephalis. The upper unpaired lobe is
located above the two smaller paired ones (Figure 3K, 3L);
these basal paired lobes are not always conspicuous. Many
pterocorythids are two or three-segmented, lacking
postabdominal segments.
Family Artostrobiidae.
Spherical or subspherical cephalis, usually with an apical
tube directed obliquely upwards (Figure 3J). The pores on all
postcephalic segments, or at least on the last ones, are
arranged in clearly defined transverse rows (Figure 3S).
Usually elongated, multisegmented forms.
Family Cannobotryidae.
Cephalis large, with several asymmetrical lobes (sometimes
appearing as irregular bulges) (Figure 3M). Mostly
2-segmented forms (cephalis and thorax), but sometimes with
post-thoracic segments.