TAXONOMY
Morphology and classification
systems
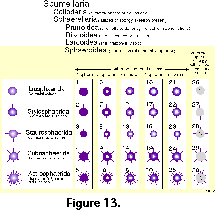 Based on his previous monograph of 1862, and
especially on the extensive collections of HMS Challenger,
Haeckel (1887) produced the first comprehensive system of
radiolarian classification, encompassing over 3,000 species,
2,400 of which were new to science. Although Haeckel's work is
still a compulsory reference guide for anyone attempting to deal
with the identification of these organisms, it has for some time
been evident that it does not satisfactorily represent natural
relationships. Indeed Haeckel’s groupings are only based on
morphological similarities without the support of continuity in
the fossil record, rather than on demonstrable evolutionary
sequences. In addition, the rigidity of his diagnoses, based
chiefly on strict geometric considerations (Figure
13), ignores the ample
intraspecific variability of the polycystines. As a result, many
of his described "species" are but slightly different
morphotypes or even developmental stages of the same organism
(see Figure 13 and Figure 14).
Based on his previous monograph of 1862, and
especially on the extensive collections of HMS Challenger,
Haeckel (1887) produced the first comprehensive system of
radiolarian classification, encompassing over 3,000 species,
2,400 of which were new to science. Although Haeckel's work is
still a compulsory reference guide for anyone attempting to deal
with the identification of these organisms, it has for some time
been evident that it does not satisfactorily represent natural
relationships. Indeed Haeckel’s groupings are only based on
morphological similarities without the support of continuity in
the fossil record, rather than on demonstrable evolutionary
sequences. In addition, the rigidity of his diagnoses, based
chiefly on strict geometric considerations (Figure
13), ignores the ample
intraspecific variability of the polycystines. As a result, many
of his described "species" are but slightly different
morphotypes or even developmental stages of the same organism
(see Figure 13 and Figure 14).
 In spite of these shortcomings and the time
elapsed, advances in the development of a better classification
system have been very limited. Efforts to depart from and improve
upon the classification schemes inherited from earlier workers
have mainly followed two different approaches: cytological
techniques and evolutionary studies.
In spite of these shortcomings and the time
elapsed, advances in the development of a better classification
system have been very limited. Efforts to depart from and improve
upon the classification schemes inherited from earlier workers
have mainly followed two different approaches: cytological
techniques and evolutionary studies.
Hollande and Enjumet
(1960), Cachon and Cachon (1972), Petrushevskaya et al.
(1976), Petrushevskaya (1981) proposed revisions which use not only the
skeleton (as most other classifications), but also cytoplasmic
features, in particular the "nucleoaxopodial complex"
(sensu Petrushevskaya 1981). Although these schemes are probably
sounder in biological terms, their applicability to fossil and
subfossil materials lacking the protoplasm is problematic, which
is one of the reasons for their very limited acceptance among
radiolarian workers.
Analyses of evolutionary lineages
in geological sequences were somewhat more succesful than
cytological techniques in defining characters applicable to
classification. Based on evolutionary evidence, Riedel and Sanfilippo
(1986) produced an
interesting critical review of the most important skeletal traits
used by Haeckel. They concluded that some of them (e.g., number
of segments, number of supplementary concentric spheres, number
of feet, number of rays and of equatorial spines in discoidal
Spumellaria, presence and nature of thoracic wings) have little
or no suprageneric value. In contrast, several others (especially
cephalic structure, but also pore arrangement, shell terminations
in Nassellaria, etc.), traditionally considered as of minor
value, are conservative through time, reveal evolutionary
lineages and, therefore, are relevant for higher-rank divisions.
Riedel (1967b, 1971),
Petrushevskaya (1965, 1971a),
Goll (1968, 1969),
Sanfilippo and Riedel
(1970), Zhamoida and Kozlova
(1971), Foreman (1973), Dumitrica (1988, 1989)
based on skeletal features alone worked out alternative
classifications, either for the entire order or for selected
polycystine groups. Of these, Riedel's (1967b, 1971)
suprageneric system has become the most widely accepted for
extant and Cenozoic radiolarians, and is the one adopted herewith
(with slight modifications; see also Kling 1978; Boltovskoy 1981e; Anderson et al. 1996). It should be stressed, however, that this
system does not overcome many of the above-mentioned problems,
and is therefore a compromise provisional classification. Several
of the family-level definitions, especially in the Spumellaria,
are rather vague and generally used as a lumping black box for
the many forms with complex morphologies and poorly understood
relationships (e.g., Litheliidae, Pyloniidae, Tholoniidae).
Specific identification of the
polycystines is a time-consuming and frustrating task. With the
exception of the few abundant and widespread species on whose
names there is fairly good agreement, binomial nomenclature alone
very often fails to pinpoint unequivocally a given morphotype
because different names are applied to the same species and,
conversely, identical organisms are reported under different
specific and even generic names (see Boltovskoy and
Jankilevich 1985). Because
a very substantial proportion of the original species
descriptions were published in old and often hard to get
monographs, some authors find it faster and easier to create a
"new species" for the unusual-looking skeleton in the
slide, than to comb the dusty books in search of an adequate,
already established name. Ecologically, paleoecologically and
stratigraphically-oriented studies often underestimate the
importance of a stable and consistent naming system; the lack of
species illustrations in these reports allows the wrong
designations to go undetected. This not only hinders buildup of
useful information, but also significantly degrades the overall
quality of radiolarian-based data for other applications. Recent
literature has abundant examples of this bias, which introduces
even more chaos into the already anarchic situation inherited
from turn of the century works. Indeed, this may be a major
reason for the waning use of radiolarians in stratigraphic and
paleoecologic work.
The following
illustrated glossary of
most commonly used terms for the description of polycystine
skeletons is chiefly based on the listing compiled by Petrushevskaya (1981). Capital letters (in parentheses) denote
the group for which the term is used (N: Nassellaria; S:
Spumellaria).
 Abdomen (N) (Figure
3O, 3P): the third
segment of nassellarian multisegmented shells.
Abdomen (N) (Figure
3O, 3P): the third
segment of nassellarian multisegmented shells.
- Aboral, aboral pole (N):
skeletal section located at the opposite extreme of the
mouth or aperture. Nassellarian growth starts with this
part of the skeleton.
- Annular strictures: see
strictures.
- Antecephalic chamber: see
antecephalic lobe.
- Antecephalic lobe (N) (Figure
3M, 3N):
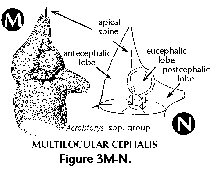 Section of the cephalis at the base
of which the dorsal spine is located. Separated from the
eucephalic lobe by the apical spine. When this section is
separated from the rest of the shell by a pored wall it
is called the antecephalic chamber.
Section of the cephalis at the base
of which the dorsal spine is located. Separated from the
eucephalic lobe by the apical spine. When this section is
separated from the rest of the shell by a pored wall it
is called the antecephalic chamber.
- Apical cupola: see galea.
- Apical horn (N) (Figure
3P): external
extension of the apical spine.
- Apical pore (N): wall-pore
located at the base of the apical spine.
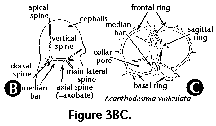
- Apical spine (N) (Figure
3A, 3B):
 Internal cephalic spine which
branches off the median bar close to the point of
insertion of the dorsal spine. The apical spine can
protrude outside of the cephalis, in which case its
external section is called apical horn.
Internal cephalic spine which
branches off the median bar close to the point of
insertion of the dorsal spine. The apical spine can
protrude outside of the cephalis, in which case its
external section is called apical horn.
- Apical tube (N) (Figure
3J): Tube-like
projection on the cephalis of the Artrostrobiidae,
homologous to the vertical spine. Also termed lateral
tube or tubule.
- Apophyses (N, S): any external
or internal protruding outgrowth of the skeletal
meshwork.
- Appendages (N): any external
or internal protruding outgrowth of the skeletal
meshwork. Riedel and Sanfilippo (1986) suggested that
primary appendages (those directly connected to or having
homologies with the internal spicule) are of higher
phylogenetic significance than the secondary ones (not
directly related to the internal spicule).
- Arches (N): anastomosed
skeletal outgrowths of the main spines, such as the one
forming the upper section of the sagittal ring (Figure
3A).
- Areolate (N, S): Referring to
small, very regularly repeated wall perforations or
pores.
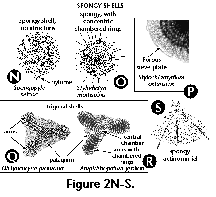
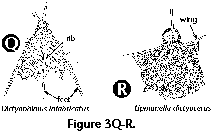
- Arms (S) (Figure
2Q, 2R): elongate
projections (usually 3) radiating from a central
subcircular disc in the shells of some Spongodiscidae.
Arms can consist of unstructured spongy meshwork (Figure
2Q), or of spongy meshwork with more or less clearly
visible chambered rings (Figure 2R).

- Axial rod: see axial spine.
- Axial spine (N) (Figure 3A,
3B): spine projecting from the median bar and oriented
toward the thorax and subsequent segments; can be simple
or branched. Also called axobate.
- Axobate: see axial spine.
- Axoneme (N, S): the central
shaft of parallel microtubules of the axopodia
(cytoplasmic structure).
- Axoplast (N, S) (Figure 1A):
central nucleus of the axopodial system (cytoplasmic
structure).
- Bars (N, S) (Figure 2F):
siliceous anastomosed beams that define the meshwork of
polycystine shells, separated by pores.
- Basal plate: see collar plate.
- Basal pores (N): see collar
pores.
- Basal ring (N) (Figure 3C):
the subcircular structure formed by the arches that
separate the wall of the cephalis from that of the
thorax. In reduced skeletons, such as those of the
Spyrida, these arches make an actual ring, whereas in
other families the ring is embedded into the shell-wall.
- Basal segment (N): the last
segment in multisegmented Nassellaria.
- Beams (S): siliceous rods
joining contiguous shell structures. In some families
(e.g., Actinommidae, Pyloniidae, Litheliidae) radial
beams connect the successive, concentric spheres (Figure
2A) or consecutive
whorls of the spiral (Figure 2L).

- Branched spines: see spines.
- Calymma (N, S) (Figure 1A):
extracapsular cytoplasm (see central capsule).
- Central capsule (N, S) (Figure
1A, 1B): an organic, perforated membrane within the
radiolarian protoplasm which separates the intracapsular
cytoplasm from the extracapsular one.
- Central chamber (S) (Figure
2R): the central, spherical structure of the skeleton of
several Spongodiscidae.
- Cephalis (N): the first
(uppermost) segment of the skeleton, which can be either
undivided (Figure 3H, 3I, 3J), or divided into lobes or
chambers (Figure 3K, 3M, 3N). The cephalis can be well
differentiated from the thorax (Figure
3F, 3H, 3I, 3J, 3K,
3L, 3M, 3N, 3R), or it can be immersed into it
partially (Figure 3D) or totally (Figure 3E).
- Cervical apophyses (N):
lateral outgrowths of the dorsal spine.
- Cervical pores (N): paired
pores in the base of the cephalis between the primary
lateral and the dorsal spines.
- Cervical stricture (N) (Figure
3O): furrow or constriction which divides the cephalis
from the thorax.
- Chamber (N): one of the
several sections into which the cephalis of the
Nassellaria can be divided (see antecephalic, lateral,
eucephalic and postcefalic chambers). See also central
chamber.
- Chambered rings (S) (Figure
2O, 2R): Concentric or spiral more or less visibly
segmented rings around the central chamber in several
Spongodiscidae (see arms).
- Club-like spines: see spines.
- Collar plate (N): the complex
structure at the base of the cephalis formed by the
spines and arches, all lying on approximately the same
plane.
- Collar pores (N) (Figure 3C):
pores in the collar plate.
- Collar stricture: see cervical
stricture.
- Columella (N): free portion of
the apical spine (Figure 3A) in the cephalis of the
Nassellaria.
- Cortical shell (S) (Figure 2A,
2B): one of the outermost perforated spherical shells of
the Spumellaria, located outside of the central capsule.
- Cupolae (S) (Figure 2C):
dome-like protuberances of the skeleton of the
Tholonidae.
- Cylindrical spines: see
spines.

- Dorsal spine (N) (Figure 3A,
3B): Internal cephalic spine projecting from the median
bar at the opposite end of the vertical spine, next to
the apical and secondary lateral spines. The dorsal spine
can protrude outside of the cephalis at the level of the
cervical stricture.
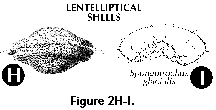
- Equatorial plane (S): mainly
applicable to lenticular, discoidal and biconvex
spumellarian shells (Figure 2H, 2I); plane of maximum shell
surface.
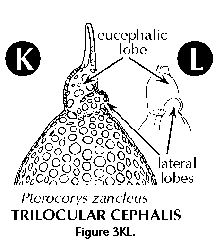
- Eucephalic lobe (N) (Figure
3N): in species with a divided cephalis, the section
which hosts the median bar.
- Extracapsular cytoplasm: see
calymma.
- Feet (N) (Figure 3Q): External
projections of the wall of the thorax oriented down and
sideways, usually in the number of 3. The feet are often
associated with the dorsal and primary lateral spines.
- First shell (S) (Figure 2A,
2B): the innermost shell of the set of concentric spheres
of an actinommid. Sometimes also applied to members of
other families with several successively larger shells
totally or partially enclosing one another. Shells are
numbered from the inside of the skeleton toward the
periphery; thus, the second shell is the one located next
to- and outside of the first one, and so on.

- Fourth shell: see first shell.
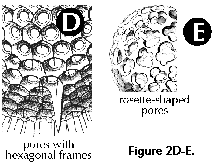
- Frames (N, S) (Figure
2D): ridges on the
surface of the shell surrounding the pores. Frames around
pores can be roundish or polygonal (pentagonal,
hexagonal, etc.).
- Fundamental spicule: see main
spicule.
- Funnel-shaped pores (N, S):
pores the external opening of which is conspicuously
wider than the internal one.
- Galea (N): helmet-shaped
portion of the shell of the Nassellaria associated with
the apical horn; its base is formed by the top of the
cephalis. Also called apical cupola.
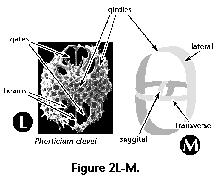
- Gates (S) (Figure 2L, M):
large openings in the skeleton's meshwork, usually
conspicuously larger than the pores (Pyloniidae).
- Girdles (S) (Figure 2L, 2M):
circular or ellipsoidal skeletal perforated plates
arranged in three mutually perpendicular planes which
form the skeleton of the Pyloniidae.
- Internal spicule: see main
spicule.
- Intracapsular cytoplasm: see
central capsule.
- Jugal pores (N): paired pores
in the base of the cephalis between the primary lateral
and the vertical spines.
- Lateral lobes (N) (Figure 3K,
3L): lateral sections of the cephalis separated from the
eucephalic portion by cephalic arches.
- Lateral spines (N) (Figure 3A,
3B): Paired spines projecting from the median bar. The
main or primary lateral spines (right and left) are
located in the vicinity of the vertical spine, while the
secondary ones (right and left) are inserted close to the
apical spine.
- Lateral tube: see apical tube.
- Latticed meshwork (N, S)
(Figure 2D, 2E, 2J, 2L): siliceous meshwork of bars
separated by regular or irregular pores of variable size,
not spongy in appearance (see spongy meshwork).

- Lentelliptical shell (S)
(Figure 2H, 2I): in the shape of a biconvex disc.
- Lumbar stricture (N) (Figure
3O): constriction between the thorax and the abdomen.
- Main lateral spines: see
lateral spines.
- Main spicule: see main spines.
- Main spines (N) (Figure 3A,
3B, 3C): basic skeletal elements of most Nassellaria,
composed of the median bar from which the apical,
vertical dorsal and lateral spines arise.
- Mantle (N, S): usually thin,
delicate, lace-like meshwork surrounding the main shell
which appears in some fully grown polycystines.
- Median bar (N) (Figure 3A, 3B,
3C): the basic nassellarian internal skeletal element
which supports the apical, vertical, dorsal and lateral
spines. Its position defines the limit between cephalis
and thorax.
- Medullary shell (S) (Figure
2A, 2B, 2J): one of the innermost perforated spherical
shells of the Spumellaria, located inside the central
capsule. The central smallest sphere (<30 µm) is also
called microsphere by some authors.
- Meshwork (N, S): the
combination of anastomosed rods and bars that form the
external siliceous skeleton of the polycystines.
- Microsphere: see medullary
shell.
- Mitral ring (N): in the
Spyrida, the skeletal ring which lies in the plane
parallel to that of the basal ring and perpendicular to
that of the sagittal ring.
- Monolocular shell (N) (Figure
15.111): Nassellarian shell composed of a single segment.
- Monothalamous shell: see
monolocular shell.
- Mouth (N): A large basal
opening in the last segment of the Nassellaria (also
called aperture), which can be open (Figure 3O, 3P) or
obliterated by a porous plate or velum.
- Multilocular cephalis (N)
(Figure 3K, 3M, 3N): cephalis provided with several
chambers or lobes in addition to the eucephalic one.
- Multilocular shell (N): see
multisegmented shell.
- Multisegmented shell (N)
(Figure 3O): Nassellarian shell composed of several (as
opposed to one) segments.
- Neck (N): elongated section of
shell joining cephalis and thorax.
- Nodes (N, S) (Figure 2F): area
of the shell where two or more bars meet (=nodal points).
- Patagium (S) (Figure 2Q, 2R):
a delicate, spongy meshwork in some Spongodiscidae which
differs from the main shell by its looser structure.
- Peristome (N) (Figure 3P):
differentiated rim around the mouth of the last segment,
usually poreless, often bearing teeth or other
structures.
- Phacoid shell (S): structure
formed by two concentric spherical medullary shells
enclosed in one lenticular cortical shell, characteristic
of the families Coccodiscidae and Phacodiscidae.
- Polar caps (S) (Figure 2J):
Additional skeletal growth in the form of latticed
cupolae at both ends of some coccodiscid shells.
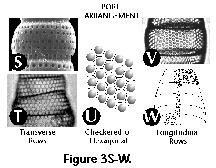
- Pore arrangement (N, S):
spatial distribution of pores of the shell-wall; regular
distributions can be checkered (Figure
3U) or in rows,
either transversal (Figure 3S, 3T) or longitudinal
(Figure 3V, 3W).
- Pores (N, S) (Figure 2D, 2E):
perforations in the skeleton of the polycystines.
- Porous sieve-plate (S) (Figure
2P): a thin, perforated plate that covers the spongy
surface of some Spongodiscidae.
- Post-abdominal (N) (Figure
3O): refers to structures associated with the fourth and
subsequent segments of multisegmented Nassellaria
(segments, pores, ribs, etc.).
- Postcephalic lobe (N) (Figure
3N): one of the sections into which the cephalis can be
divided, sometimes elongated into a tube.
- Primary lateral spines: see
lateral spines.
- Primary spines: see spines.
- Pylome (S) (Figure 2N): A
larger opening on the external shell of some Spumellaria,
often extending into a tube or surrounded by larger
spines.
- Ribs (N) (Figure 3Q):
elongated, usually unperforated thickenings in the
shell-wall. Sometimes the ribs can extend into wings
(Figure 3R) or feet (Figure 3Q). Dorsal and primary
lateral spines embedded in the wall of the thorax can
also form ribs.
- Rosette-shaped (N, S): usually
applicable to pores with a tri- or tetra-lobed outline
(Figure 2E).
- Sagittal plane (N): the plane
defined by the position of the median bar (and also the
apical, vertical, dorsal and axial spines).
- Sagittal ring (N) (Figure 3A):
a skeletal ring formed by the median bar at the base, the
vertical and apical spines at the front and back,
respectively, and an arch joining these two spines at the
top. It can be either free or embedded into the skeletal
meshwork.
- Second shell: see first shell.
- Secondary lateral spines: see
lateral spines.
- Secondary spines (=
by-spines): see spines.
- Segment (N) (Figure 3O): one
of the several joints or sections which compose a
Nassellarian skeleton, separated by the adjoining ones by
an internal circular thickening of the shell-wall and/or
by an external stricture.
- Septae (N): Well-developed
internal divisions between the segments of nassellarian
shells.
- Sieve-plate: see porous
sieve-plate.
- Spines (N, S): any internal or
external rod or needle with one free end. According to
their relative size and number, spumellarian spines can
be primary (larger, usually fewer in number; Figure 2B)
and secondary (smaller, more numerous, sometimes bristle-
or thorn-shaped; Figure 2B). In cross-section spines can
be triangular (three-bladed) with smooth or serrated
edges, or cylindrical; straight, or twisted (Figure 2G).
The distal end of spines can be unbranched or branched
(Figure 2G). Club-like spines have a swollen or thickened
distal end (Figure 2G). Radial spines are those radiating
centrifugally from the center of the shell (Figure 2B).
- Spongy meshwork (N, S) (Figure
2N, 2O, 2P, 2Q, 2R, 2S): sponge-like (as opposed to
latticed) siliceous meshwork of thin, irregular, more or
less densely packed bars, mainly in some spumellarian
families (Actinommidae, Spongodiscidae).
- Strictures (N) (Figure 3O):
joints between contiguous segments, often marked by
conspicuous external constrictions.
- Teeth (N) (Figure 3P):
protruding apophyses around the mouth, of variable length
and thickness (=oral teeth, adoral teeth, terminal teeth,
subterminal teeth).
- Terminal teeth: see teeth.
- Third shell: see first shell.
- Thorax (N) (Figure 3O, 3P):
second segment of multisegmented nassellarians.
- Tripod (N): section of the
internal skeleton formed by the dorsal and primary
lateral spines (Figure 3A, 3B). In some shells the tripod
protrudes externally forming the feet (Figure 3Q).
- Tubercles (N, S): pustule-like
lumps (condyles, mamillae) on the surface of the
siliceous shell.
- Unilocular shell: see
monolocular shell.
- Velum (N): a porous plate
obliterating the mouth.
- Ventral spine: same as axial
spine.
- Vertical spine (N) (Figure 3A,
3B): Main unpaired spine projecting from the median bar
in the vicinity of the primary lateral spines and the
axial spine.
- Wings (N) (Figure 3R): lateral
apophyses of the thorax usually connected to the
shell-wall by siliceous meshwork, derived from external
projections of the dorsal and primary lateral spines.