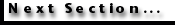3. THE REALISATION OF NEPTUNE - THE REAL WORLD IS WORSE
THAN WE THOUGHT
3.1. Stratigraphic and geographic coverage
The geographic distribution of the 165 holes included in Neptune is uneven.
In some areas there is a very detailed coverage, like for example in some parts
of the Antarctic Ocean (Fig. 2.1). On
the other hand, no holes from the central north and southeastern Pacific Ocean
are present in Neptune. (Holes from ODP Leg 145 now provide a transect across
the north Pacific.) The mid- and high latitudes in the southern hemisphere and
the tropical regions of the Atlantic Ocean are also not well represented. This
is due in part to the uneven coverage of DSDP and ODP cruises and in part to the
selection made for Neptune, which preferentially included holes with good
biostratigraphy.
Each year, ODP organizes five to six drilling cruises which result in as many
published Scientific Results. Although not all cruises retrieve
micropaleontologically significant material, many of them provide a detailed
biostratigraphy and data relevant to the scopes of Neptune. The present
geographic coverage of holes in Neptune has been last updated in 1995 (Leg 135).
Since then more than twenty-five volumes of Scientific Results have been
published. From the beginning of the project, we were faced with the need to
maintain a balance between keeping up with the new data produced by ODP and the
need to analyze the data already in Neptune for biostratigraphic or
micropaleontological studies. At present, I have decided to keep Neptune at its
current, acceptable but not optimal, size in order to complete some of the
studies that we had planned. If it will be decided to update Neptune in the
future, it will be necessary to:
- select suitable holes from Leg 136 onward, and for the selected holes:
- compile biostratigraphic files and construct age models;
- import the core depth file and the age model file for each hole;
- download from ODP the available range charts;
- format the range chart files to make them compatible with Neptune;
- import the range chart file;
- update the species name list with the new names eventually present in
the range charts.
 Another
limitation of the database is given by the often incomplete often-incomplete
recovery of sediments (Fig. 3.1).
Before the advent of hydraulic piston coring, few continuously recovered
sections were available. Core recovery has drastically improved in the more
recent ODP holes but sediment loss at core breaks is still common even in
continuously cored sections (Farrell
and Janecek 1991).
Another
limitation of the database is given by the often incomplete often-incomplete
recovery of sediments (Fig. 3.1).
Before the advent of hydraulic piston coring, few continuously recovered
sections were available. Core recovery has drastically improved in the more
recent ODP holes but sediment loss at core breaks is still common even in
continuously cored sections (Farrell
and Janecek 1991).
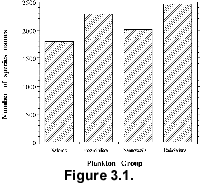
In addition, there is an uneven distribution in the temporal coverage of the
sections. Whilst Plio-Pleistocene sections are very well represented in Neptune
(as they are in ODP holes overall), the detail of stratigraphic coverage
decreases for older time periods, as naturally expected from the drilling
procedure (Fig. 3.2, Spencer-Cervato
1998). This might be interpreted as a need to recover more Miocene and older
sections, but this pattern actually reflects the number of studied
sections and not simply the recovered sections. 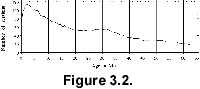 Therefore,
I believe that the problem does not lie only in the quantity of older
sections drilled, but also in the quality of their stratigraphy. The
reliability of the stratigraphy provided for a section depends strongly on the
availability of good calibrations, and these are currently available mainly for
Neogene sediments. Figure 3.2 also
shows that the number of well-studied sections does not decrease gradually and
regularly with age, but shows peaks (around 2 Ma) and plateaus (e.g., between 20
and 32 Ma). This likely reflects the relative, unequal attention given to the
Cenozoic stratigraphy through the history of DSDP and ODP.
Therefore,
I believe that the problem does not lie only in the quantity of older
sections drilled, but also in the quality of their stratigraphy. The
reliability of the stratigraphy provided for a section depends strongly on the
availability of good calibrations, and these are currently available mainly for
Neogene sediments. Figure 3.2 also
shows that the number of well-studied sections does not decrease gradually and
regularly with age, but shows peaks (around 2 Ma) and plateaus (e.g., between 20
and 32 Ma). This likely reflects the relative, unequal attention given to the
Cenozoic stratigraphy through the history of DSDP and ODP.
3.2. Chronology
For the database, we have chosen to use a comprehensive biochronology based
on deep-sea sections, therefore not considering land sections, which represent
the type localities where stratigraphic series were first described. This may
represent a limitation in the achieved biochronological calibration. The
precision of the ages determined with the age models depends on various factors,
some subjective and nonquantifiable, and some, like sample spacing, accuracy of
biostratigraphic calibration, or core recovery, that can be quantified. A
conservative estimate of the age model precision of 0.36 m.y. was determined for
Neogene sediments (Spencer-Cervato
et al. 1994). For Paleogene sediments it is about 0.66 m.y. (twice the
average sample spacing).
Another important factor is the quality of the age model. The Neogene DSDP
age/depth plots that we have published so far (Spencer-Cervato
et al. 1993; Lazarus
et al. 1995a) are a good example of the range of reliability of the line of
correlation. The subjective ranking given in Table
2.2 varies from very poor or poor (wide scatter of events, straight line of
correlation drawn across the middle of the cloud), to moderate (some scatter of
a limited number of events, various possible lines of correlation), to good or
excellent (40% of the holes: very good agreement of the event ages, abundant
events to constrain the line of correlation, good agreement between
magnetostratigraphy and biostratigraphy). Several factors can cause the scatter
of events observed in most age/depth plots: reworking, downhole contamination,
incorrect entry in the biostratigraphy file, typos in the range charts,
diachrony of the calibrated event, , and sample spacing. Whilst most of these
causes can be easily double-checked, diachrony is quite difficult to assess. The
assumption of globally synchronous events which is at the base of
biochronology is validly established only for some selected, well documented
events (e.g., Hays
and Shackleton 1976; Thierstein
et al. 1977; Backman and
Shackleton 1983; Wei
1993; Spencer-Cervato
et al. 1994). It is likely that more complete data collection and
documentation would lead to the identification of more globally synchronous
events. But in most cases, a calibration is valid only for the more or less
restricted biogeographic province where it is done, and only a few events are
truly globally synchronous, within the precision of the method adopted for
calibration. The need for localized calibrations has long been known for
siliceous plankton stratigraphy, but it is not widely accepted by
biostratigraphers using calcareous plankton. To minimize this factor, we
intentionally used multiple regional calibrations for diatoms and radiolarians.
Even with this approach, the scatter is sometimes too large to provide a
reliable line of correlation. For nannofossils and foraminifera only one general
(low latitude) calibration is available (Berggren
et al. 1985, 1995a,
b). The advantage
of this calibration is that it is based on several sites, while most of the
regional calibrations are based only on one hole. An estimate of the diachrony/synchrony
of Neogene events was done with a subset of the holes currently present in
Neptune (Spencer-Cervato
et al. 1994). This study indicated that calcareous nannofossils provide the
most reliable biostratigraphic events, as they are mostly cosmopolitan and, if
diachronous, the age margin is relatively small.
 Very
few sections are actually continuous, and long stratigraphic gaps are common (Spencer-Cervato
1998) (Fig. 3.3). Two-thirds of the
selected holes contain at least one hiatus, and on average they each contain
three hiati of various lengths (Fig. 2.1).
The presence of these hiati results in artificially older or younger ages for
the samples adjacent to the gap. This does not allow one to automatically (blindly)
search the database for e.g.,such information as species ages, but requires that
every output is be checked and compared with the age models.
Very
few sections are actually continuous, and long stratigraphic gaps are common (Spencer-Cervato
1998) (Fig. 3.3). Two-thirds of the
selected holes contain at least one hiatus, and on average they each contain
three hiati of various lengths (Fig. 2.1).
The presence of these hiati results in artificially older or younger ages for
the samples adjacent to the gap. This does not allow one to automatically (blindly)
search the database for e.g.,such information as species ages, but requires that
every output is be checked and compared with the age models.
The final and probably most necessary improvement of the chronology of
Neptune is given by the life-timelifetime of the biochronology selected for the
age model calibration. We initially based the age models on Berggren
et al. (1985). An updated magnetostratigraphy was published later (Cande and
Kent 1992) but it did not provide the combination of biochronology and
magnetostratigraphy available from Berggren
et al.'s (1985) work. We thus decided to continue using Berggren
et al. (1985) throughout the first phase of the project (DSDP Neogene
sediments). However, ten years after the first biochronology compilation, a new
updated biochronology was published (Berggren
et al. 1995b) and the chronology of Neptune became suddenly outdated. The
iterations to update Neptunes chronology were greatly helped by additional
programming of Neptune by Dave Lazarus and an auxiliary computer program (not
part of Neptune and written ad hoc by Bernhard Brabec) which created a
correlation function between the old and the new master biochronology. This
function was applied to all age model files and new revised age models were
created. . Then, all biostratigraphy files were updated using a find -
replace routine with lookup tables (i.e.: if code in column 3 is equal to
xYwz, replace age in column 4 with corresponding value in lookup table). While
we could directly use the new calibrations for calcareous plankton as lookup
tables, it was necessary to recalibrate to the new time scale all regional
templates used for siliceous plankton events. Finally, before the new age models
could be imported into Neptune, all the age/depth plots were redone by myself
and eventually adjusted to fit the new event ages.
Among the other reasons mentioned above, if a bio-event recorded in a
specific hole plots far outside the area where the line of correlation can be
drawn, it could be due to its taxonomic identification. Many authors have put
together the hundreds of range charts that were used for Neptune and not all
agree in the detailed taxonomic identification of all the 8800+ taxa included in
Neptune. Indeed, taxonomic identification is subjective. The time pressure under
which biostratigraphers are during a leg is also an important limiting factor in
the number of species described in a range chart, which is often limited to
biostratigraphic markers. The extent to which this taxonomic problem has
affected the data in Neptune can be judged by experts in particular cases but
cannot be easily quantified.
Starting from the biostratigraphy filesbiostratigraphic records assembled for
the chronology, we assumed that the taxon associated with one event and
described in the range chart was the one we were looking for. Further, we needed
to consider the occurrence of synonyms. It sometimes happens that the name used
by one author for a taxon corresponds either to a different taxon according to
another author, or that a different name is used by a second author for this
specific taxon (synonymy). For example, the foraminifer species Globorotalia
truncatulinoides has been also called Truncorotalia truncatulinoides.
To account for this, we have used the literature, personal experience and
extensive consultation with taxonomic experts to identify valid taxon names. 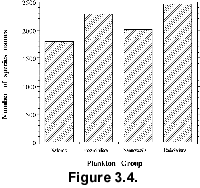 Three
thousand of the 8810 names listed in Neptune (Fig.
3.4) are considered valid (i.e., are legal names in the framework of the
ICZN and ICBN, and are known to be real to at least one of the experts).
Synonyms to these valid names were then identified (with the corresponding valid
name). They constitute 31% of the total number of names. In several cases we
could not unequivocally identify a specific name and marked it as unknown
(15% of all names). Only 43 names (0.5%) were considered invalid. This
information is available in the Species Names table of Neptune. The
synonymizsation is subjective (the initials of the person who identified each
species name is also given in the Species Names table) and the names list
does not at all pretend to be a thorough or complete taxonomic revision of
marine plankton. It merely represents a working table that gives us a first
approximation of plankton taxonomy. A real taxonomic database would need
complete taxonomic descriptions (with history) for each taxon and a series of
images to illustrate them. Cathy Nigrini, Jean-Pierre Caulet, and Dave Lazarus
are currently working on a detailed taxonomic database for radiolarians, but it
is well beyond the scopes of Neptune to even attempt anything like this for all
groups.The taxonomic list also needs continuous update: every time a new hole is
added to Neptune, the biostratigraphic range charts carry with them new names,
sometimes several ones. These need to be added to the Species Names list
and identified as valid or not.
Three
thousand of the 8810 names listed in Neptune (Fig.
3.4) are considered valid (i.e., are legal names in the framework of the
ICZN and ICBN, and are known to be real to at least one of the experts).
Synonyms to these valid names were then identified (with the corresponding valid
name). They constitute 31% of the total number of names. In several cases we
could not unequivocally identify a specific name and marked it as unknown
(15% of all names). Only 43 names (0.5%) were considered invalid. This
information is available in the Species Names table of Neptune. The
synonymizsation is subjective (the initials of the person who identified each
species name is also given in the Species Names table) and the names list
does not at all pretend to be a thorough or complete taxonomic revision of
marine plankton. It merely represents a working table that gives us a first
approximation of plankton taxonomy. A real taxonomic database would need
complete taxonomic descriptions (with history) for each taxon and a series of
images to illustrate them. Cathy Nigrini, Jean-Pierre Caulet, and Dave Lazarus
are currently working on a detailed taxonomic database for radiolarians, but it
is well beyond the scopes of Neptune to even attempt anything like this for all
groups.The taxonomic list also needs continuous update: every time a new hole is
added to Neptune, the biostratigraphic range charts carry with them new names,
sometimes several ones. These need to be added to the Species Names list
and identified as valid or not.
3.4. Biostratigraphy
There is an uneven distribution in the number of reports by plankton group in
Neptune. Over 60% of the 225 articles from which data for Neptune have been
extracted (Table 2.4) are on calcareous
plankton, almost equally distributed between nannofossils and foraminifera. 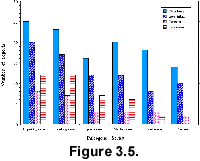 Radiolarians
follow with about 21% and diatoms trail with only 16%. At the same time,
biostratigraphic work on siliceous plankton is underrepresented in Paleogene
sections, and most often limited to the Oligocene and younger sections (Fig.
3.5). This unevenness represents a bias for evolution studies where we would
like to compare calcareous and siliceous plankton occurrences. Whether this
distribution represents the average abundance of fossil plankton in deep-sea
deposits or is instead the reflection of staffing decisions by DSDP and ODP is
yet to be determined.
Radiolarians
follow with about 21% and diatoms trail with only 16%. At the same time,
biostratigraphic work on siliceous plankton is underrepresented in Paleogene
sections, and most often limited to the Oligocene and younger sections (Fig.
3.5). This unevenness represents a bias for evolution studies where we would
like to compare calcareous and siliceous plankton occurrences. Whether this
distribution represents the average abundance of fossil plankton in deep-sea
deposits or is instead the reflection of staffing decisions by DSDP and ODP is
yet to be determined.
One of the limitations of Neptune as a comprehensive micropaleontological
database is given by our decision to include only four plankton groups. The DSDP
and ODP Reports include many articles on benthic foraminifera, silicoflagellates,
dinoflagellates as well as palynology. At the moment, there are no plans to
include their occurrence data in the database, which in itself would not be a
huge task.