There are several similarities in the isotopic
patterns of Giganotosaurus and Tyrannosaurus with one major
difference. In both dinosaurs, the vertebrae are the warmest (lowest ![]() 18O
values) bones sampled. The cervicals and dorsal from Giganotosaurus
centra show less variability than in the T. rex specimen. Previously, the
ribs and dorsal vertebrae were considered as "core" bones
representative of the warmest body temperatures. However, the mid section and
distal end of ribs are in the outer shell of the body and may represent
temperatures more similar to those of the extremities than the core body. In T.
rex, values from the gastralia were similar to those from the femur (Barrick
and Showers 1994). In both T. rex
and Giganotosaurus, the ribs are 2-4°C cooler than the vertebra. The Giganotosaurus
gastralia do have greater intrabone variability than in the T. rex individual.
Both individuals display regional heterothermy in the limbs with cooler
temperatures and greater isotopic variability occurring with greater distance
from the core body. In both cases the femora are 3-4°C cooler than the
vertebrae and the tibia 4-5°C cooler than the vertebrae. Only in Giganotosaurus
were proximal and distal ends of these bones compared. No difference in mean
temperature or variability was found along the femur while the tibia exhibited a
trend of increasing temperature variability (1°C) and cooler mean temperature
(1°C) distally along the shaft. Unfortunately there were no pedal elements
preserved in Giganotosaurus. The consistency in trends of increasing
isotopic variability and decreasing mean temperature distally along the limb
elements between the Giganotosaurus and Tyrannosaurus suggest that
the foot in Giganotosaurus was likely cooler and more variable than the
tibia. Thus, the
18O
values) bones sampled. The cervicals and dorsal from Giganotosaurus
centra show less variability than in the T. rex specimen. Previously, the
ribs and dorsal vertebrae were considered as "core" bones
representative of the warmest body temperatures. However, the mid section and
distal end of ribs are in the outer shell of the body and may represent
temperatures more similar to those of the extremities than the core body. In T.
rex, values from the gastralia were similar to those from the femur (Barrick
and Showers 1994). In both T. rex
and Giganotosaurus, the ribs are 2-4°C cooler than the vertebra. The Giganotosaurus
gastralia do have greater intrabone variability than in the T. rex individual.
Both individuals display regional heterothermy in the limbs with cooler
temperatures and greater isotopic variability occurring with greater distance
from the core body. In both cases the femora are 3-4°C cooler than the
vertebrae and the tibia 4-5°C cooler than the vertebrae. Only in Giganotosaurus
were proximal and distal ends of these bones compared. No difference in mean
temperature or variability was found along the femur while the tibia exhibited a
trend of increasing temperature variability (1°C) and cooler mean temperature
(1°C) distally along the shaft. Unfortunately there were no pedal elements
preserved in Giganotosaurus. The consistency in trends of increasing
isotopic variability and decreasing mean temperature distally along the limb
elements between the Giganotosaurus and Tyrannosaurus suggest that
the foot in Giganotosaurus was likely cooler and more variable than the
tibia. Thus, the ![]() 18O
values between the dorsal/cervical vertebrae, ribs and limbs of both animals are
strikingly similar suggesting similar thermoregulatory patterns. However, the
18O
values between the dorsal/cervical vertebrae, ribs and limbs of both animals are
strikingly similar suggesting similar thermoregulatory patterns. However, the ![]() 18O
values in the caudal vertebrae differ significantly. In T. rex, the
caudal vertebrae exhibit similar intrabone patterns and gradually become more
positive (~0.5-4°C cooler) with distance from the torso (Barrick
and Showers 1994). In Giganotosaurus,
however the caudal vertebrae exhibit the same mean temperature values as the
dorsal and cervical vertebrae. The mean values from the proximal caudals
resembled those of the dorsal vertebrae in both individuals. Yet in T. rex
intrabone temperature variability in the 3 caudals analyzed (~2°C) was less
than that in 4 Giganotosaurus caudals ranging from 2.5-5.5°C.
18O
values in the caudal vertebrae differ significantly. In T. rex, the
caudal vertebrae exhibit similar intrabone patterns and gradually become more
positive (~0.5-4°C cooler) with distance from the torso (Barrick
and Showers 1994). In Giganotosaurus,
however the caudal vertebrae exhibit the same mean temperature values as the
dorsal and cervical vertebrae. The mean values from the proximal caudals
resembled those of the dorsal vertebrae in both individuals. Yet in T. rex
intrabone temperature variability in the 3 caudals analyzed (~2°C) was less
than that in 4 Giganotosaurus caudals ranging from 2.5-5.5°C.
The features indicative of endothermy (defined as the maintenance a high and constant body temperature by metabolic means by Bennett and Ruben 1979) in Giganotosaurus include the homeothermy of the cervical and dorsal vertebra and regional heterothermy of the limbs. The pattern is even more suggestive than for T. rex as the core (vertebral) temperature range is nearly 1.5°C narrower in Giganotosaurus. Clarke and Jenkyns (1999) report maximum Albian Cenomanian sea-surface temperatures of 16-18°C for southern latitudes of 49°-53°S. This strongly suggests that the terrestrial climate was temperate where Giganotosaurus roamed in the Neuquen basin. Such a climate provides evidence against the warm homogenous environment in which gigantothermy could provide stable core body temperatures. In order to maintain the thermoregulatory pattern seen in Giganototsaurus, an elevated metabolic rate is required. However, the pattern of high intrabone variability and no mean interbone differences in the tail is also typical of a mass homeotherm (Barrick et al. 1997). Core homeothermy and regional limb heterothermy indicate that both individuals maintained metabolic levels above those of modern reptiles. While core homeothermy within a temperate climate fits the definition of endothermy it is likely that these dinosaurs possessed intermediate metabolic levels rather than modern mammalian levels. A drop in mass-specific metabolic rates between juveniles and adults at this intermediate level would also explain the mass homeotherm-like isotopic pattern seen in the Giganotosaurus tail. In adults, bone turnover rates in vertebrae are much more rapid than in limb bones (Francillon-Vieillot et al. 1990) and reflect a narrower time of bone deposition. Thus, a change in thermoregulatory style would be presented in the vertebrae more rapidly than in the limbs. Endothermy has come to be synonymous with mammalian and avian tachymetabolism where metabolic rates are 10x greater than for ectotherms (e.g., Ruben 1995). Endogenous heat production through elevated metabolic rates is required for endothermy, but intermediate levels (e.g. 5x metabolic rate of ectotherms) may have been high enough for the maintenance of homeothermy in dinosaurs, thus making them at least intermediate endotherms.
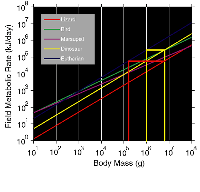 At
an adult body mass of 8000 kg, even if Giganotosaurus began life with a
modern avian metabolic level, it would have had an intermediate metabolic rate
the equivalent of a 1000 kg mammalian carnivore. At 6000 kg, the metabolic rate
of T. rex would have been equivalent to an 800 kg mammal. The dinosaurian
metabolic rate is calculated by projecting the mass vs. field metabolic rate
regression of modern avians, mammals, and reptilians (Nagy
1987) to a mass of 6000 and 8000 kg (Fig.
4) and projecting a hypothesized dinosaur regression midway between the
reptilian and eutherian mammal regression lines. In fact, at this weight (~7000
kg), the avian regression line falls midway between the projected eutherian
mammal and reptilian regression lines, intersecting the hypothesized
intermediate dinosaurian regression line. Theropods are more closely related to
birds (Ji-Qiang
et al. 1998) than other dinosaurs and a
projected intermediate metabolic level for 6000-8000 kg birds, fits well with
the interpretation of intermediate metabolic levels for these large theropods
based on their
At
an adult body mass of 8000 kg, even if Giganotosaurus began life with a
modern avian metabolic level, it would have had an intermediate metabolic rate
the equivalent of a 1000 kg mammalian carnivore. At 6000 kg, the metabolic rate
of T. rex would have been equivalent to an 800 kg mammal. The dinosaurian
metabolic rate is calculated by projecting the mass vs. field metabolic rate
regression of modern avians, mammals, and reptilians (Nagy
1987) to a mass of 6000 and 8000 kg (Fig.
4) and projecting a hypothesized dinosaur regression midway between the
reptilian and eutherian mammal regression lines. In fact, at this weight (~7000
kg), the avian regression line falls midway between the projected eutherian
mammal and reptilian regression lines, intersecting the hypothesized
intermediate dinosaurian regression line. Theropods are more closely related to
birds (Ji-Qiang
et al. 1998) than other dinosaurs and a
projected intermediate metabolic level for 6000-8000 kg birds, fits well with
the interpretation of intermediate metabolic levels for these large theropods
based on their ![]() p
distribution. The thermoregulatory pattern in Giganotosaurus corroborates
the interpretation based on the pattern seen in T. rex. If we assume that
all adults of Giganotosaurus and Tyrannosaurus reflect
the same thermoregulatory patterns as the two individuals sampled, they would
have enjoyed the benefits of endothermy at metabolic levels intermediate between
those of modern mammals and reptilians. An extension of this interpretation is
that juvenile theropods also maintained intermediate if not high metabolic
levels. Comparatively, an 8000 kg lizard or crocodile would have the same
metabolic food requirements as a 200 kg carnivorous mammal (i.e., medium sized
male African lion Panthera leo). Food requirements for various sized
eutherian mammals can be calculated from the generalized equation y=axb where
y is the food requirement in g/day, a=0.235, x is the mass of the individual in
g, and b=0.822 (eq. 19 in Nagy
1987). By converting dinosaurian or
reptilian metabolic rates to equivalent mammalian metabolic rates this single
equation can be used to estimate food requirements for these dinosaurs. Using
the intermediate metabolic rates for Giganotosaurus (8000 kg) and Tyrannosaurus
(6000 kg), their food requirements would have been 20 and 17 kg/day
respectively. This is the same food requirement for 3-4 large male tigers or
African lions. Modern male lions (Panthera leo) reach only ~250 kg while
large male tigers (Panthera tigris) may reach just over 300 kg (Walkers
Mammals of the World, 1991). For
comparative purposes, had Giganotosaurus maintained a strictly reptilian
metabolic rate, it would have required 5.5 kg/day, whereas a Giganotosaurus
individual with a eutherian metabolic rate would have required 111 kg/day of
food. Thus, for under 4x rather than 20x the food requirements of a similarly
sized bradymetabolic ectotherm or mass homeotherm, Giganotosaurus enjoyed
the benefits of endothermy. These benefits include, core homeothermy 365 days a
year, higher activity levels, and rapid growth rates. Concomitantly, these
dinosaurs would also have kept some of the benefits typical of bradymetabolic
ectotherms (e.g., lower food requirements, greater percentage of energy budget
directed toward growth and reproduction). Table 1 [what is this???]
records the food requirements of a tyrannosaur or giganotosaur at various body
sizes throughout ontogeny assuming this intermediate metabolic rate. Each adult
tyrannosaur or giganotosaur would have required the equivalent of 2 large
ornithopods or 1 subadult sauropod (~7000 kg) per year. The amount of food
required to sustain a metabolism high enough to maintain core homeothermy in
these animals is much smaller than their stomach volume. Thus, these dinosaurs
could have eaten much more than required for maintenance of their metabolism. A
large male African lion (Panthera leo) weighing 250 kg may eat up to 40
kg of meat in one meal (Walkers
Mammals of the World, 1991) which is
enough meat to support it for 7-8 days. 6000-8000 kg theropods could have held
10 times this amount of food in their stomachs. Thus, these dinosaurs could have
stored 2-3 weeks food requirements in their stomachs with a single large meal. A
truly bradymetabolic tyrannosaur or giganotosaur needing but 2000 kg of meat per
year could have survived on 5 good meals/year. Evidence of relatively unaltered
bone in a tyrannosaur coprolite indicates that residence time of food in these
theropod digestive tracts was short (Chin
et al. 1998; Bartlett
et al. 1998) suggesting that these
dinosaurs ate more often than once every 2-3 weeks. Evidence of soft tissue in
dinosaur coprolites suggest even shorter residence times (Bertrand
1903; Chin
1999). Greater food intake would have
supported increased activity levels and or growth rates. An intermediate
metabolic level has important implications for growing juvenile theropods. For
example, 100 to 1000 kg growing theropods would have needed 1-6 kg of food/day
while 2000-5000 kg individuals would have needed 10-16 kg/day. Much greater
amounts of food would have been available to these individuals and could have
been held in their digestive systems. Energy from greater food acquisition
greater than required by field metabolism could be put directly into growth, as
the maintenance of homeothermy would have allowed continuous annual growth.
Thus, greater amounts of energy were available for growth in these theropods
than is available for mammalian style endotherms. More continuous, rapid growth
due to maintenance of homeothermy would have been available for these dinosaurs
than is available for bradymetabolic ectotherms. Curry
(1998) and Ricqles
et al. (1998) have suggested extremely
rapid growth rates for Apatosaurus (=75% adult size in 7-10 yrs) and Maiasaura
(adult size in 8 yrs or less) based upon histological analyses. Intermediate
metabolic rates suggested here, allowing the maintenance of core homeothermy at
all sizes, would have supported similarly rapid growth rates for these theropods
as is suggested for Apatosaurus and Maiasaura. Intermediate
metabolic rates are also consistent with the isotopic patterns of several
herbivorous dinosaurs (Barrick
et al. 1996, 1998).
On the other hand, this intermediate metabolic strategy would have these
individuals to survive longer periods during resource scarcity than true modern
endotherms
p
distribution. The thermoregulatory pattern in Giganotosaurus corroborates
the interpretation based on the pattern seen in T. rex. If we assume that
all adults of Giganotosaurus and Tyrannosaurus reflect
the same thermoregulatory patterns as the two individuals sampled, they would
have enjoyed the benefits of endothermy at metabolic levels intermediate between
those of modern mammals and reptilians. An extension of this interpretation is
that juvenile theropods also maintained intermediate if not high metabolic
levels. Comparatively, an 8000 kg lizard or crocodile would have the same
metabolic food requirements as a 200 kg carnivorous mammal (i.e., medium sized
male African lion Panthera leo). Food requirements for various sized
eutherian mammals can be calculated from the generalized equation y=axb where
y is the food requirement in g/day, a=0.235, x is the mass of the individual in
g, and b=0.822 (eq. 19 in Nagy
1987). By converting dinosaurian or
reptilian metabolic rates to equivalent mammalian metabolic rates this single
equation can be used to estimate food requirements for these dinosaurs. Using
the intermediate metabolic rates for Giganotosaurus (8000 kg) and Tyrannosaurus
(6000 kg), their food requirements would have been 20 and 17 kg/day
respectively. This is the same food requirement for 3-4 large male tigers or
African lions. Modern male lions (Panthera leo) reach only ~250 kg while
large male tigers (Panthera tigris) may reach just over 300 kg (Walkers
Mammals of the World, 1991). For
comparative purposes, had Giganotosaurus maintained a strictly reptilian
metabolic rate, it would have required 5.5 kg/day, whereas a Giganotosaurus
individual with a eutherian metabolic rate would have required 111 kg/day of
food. Thus, for under 4x rather than 20x the food requirements of a similarly
sized bradymetabolic ectotherm or mass homeotherm, Giganotosaurus enjoyed
the benefits of endothermy. These benefits include, core homeothermy 365 days a
year, higher activity levels, and rapid growth rates. Concomitantly, these
dinosaurs would also have kept some of the benefits typical of bradymetabolic
ectotherms (e.g., lower food requirements, greater percentage of energy budget
directed toward growth and reproduction). Table 1 [what is this???]
records the food requirements of a tyrannosaur or giganotosaur at various body
sizes throughout ontogeny assuming this intermediate metabolic rate. Each adult
tyrannosaur or giganotosaur would have required the equivalent of 2 large
ornithopods or 1 subadult sauropod (~7000 kg) per year. The amount of food
required to sustain a metabolism high enough to maintain core homeothermy in
these animals is much smaller than their stomach volume. Thus, these dinosaurs
could have eaten much more than required for maintenance of their metabolism. A
large male African lion (Panthera leo) weighing 250 kg may eat up to 40
kg of meat in one meal (Walkers
Mammals of the World, 1991) which is
enough meat to support it for 7-8 days. 6000-8000 kg theropods could have held
10 times this amount of food in their stomachs. Thus, these dinosaurs could have
stored 2-3 weeks food requirements in their stomachs with a single large meal. A
truly bradymetabolic tyrannosaur or giganotosaur needing but 2000 kg of meat per
year could have survived on 5 good meals/year. Evidence of relatively unaltered
bone in a tyrannosaur coprolite indicates that residence time of food in these
theropod digestive tracts was short (Chin
et al. 1998; Bartlett
et al. 1998) suggesting that these
dinosaurs ate more often than once every 2-3 weeks. Evidence of soft tissue in
dinosaur coprolites suggest even shorter residence times (Bertrand
1903; Chin
1999). Greater food intake would have
supported increased activity levels and or growth rates. An intermediate
metabolic level has important implications for growing juvenile theropods. For
example, 100 to 1000 kg growing theropods would have needed 1-6 kg of food/day
while 2000-5000 kg individuals would have needed 10-16 kg/day. Much greater
amounts of food would have been available to these individuals and could have
been held in their digestive systems. Energy from greater food acquisition
greater than required by field metabolism could be put directly into growth, as
the maintenance of homeothermy would have allowed continuous annual growth.
Thus, greater amounts of energy were available for growth in these theropods
than is available for mammalian style endotherms. More continuous, rapid growth
due to maintenance of homeothermy would have been available for these dinosaurs
than is available for bradymetabolic ectotherms. Curry
(1998) and Ricqles
et al. (1998) have suggested extremely
rapid growth rates for Apatosaurus (=75% adult size in 7-10 yrs) and Maiasaura
(adult size in 8 yrs or less) based upon histological analyses. Intermediate
metabolic rates suggested here, allowing the maintenance of core homeothermy at
all sizes, would have supported similarly rapid growth rates for these theropods
as is suggested for Apatosaurus and Maiasaura. Intermediate
metabolic rates are also consistent with the isotopic patterns of several
herbivorous dinosaurs (Barrick
et al. 1996, 1998).
On the other hand, this intermediate metabolic strategy would have these
individuals to survive longer periods during resource scarcity than true modern
endotherms
![]()