APPENDICULAR MORPHOLOGYAs is the case for most Santa Cruz Formation mammals, descriptions and detailed illustrations of the postcrania of Protypotherium were published in the Reports of the Princeton University Expedition to Patagonia, 1896-1899 (see Sinclair 1909). We include relevant illustrations here and add functional interpretations to Sinclair's physical descriptions. Some of the volumes of this series are available free at the Internet Archive. The scapula is roughly quadrangular in overall shape; the spine runs nearly parallel to the caudal border, and intersects the superior (vertebral) border at an angle close to 90°. The supraspinous fossa is larger than the infraspinous fossa. Although a small extension for the attachment of teres major is present, it is less developed than in fossorial taxa (Shockey et al. 2007). The scapula contrasts markedly with the triangular scapulae of artiodactyls and some rodents, more closely resembling those of canids and the generalized mammal Didelphis. The unbroken length of the metacromion process in Protypotherium is unknown. The presence of a long, slender metacromion has been correlated with quadrupedal bounding in modern mammals such as lagomorphs (Janis and Seckel 2005), but the shape of this structure in Protypotherium does not closely resemble that seen in modern lagomorphs. Metacromia of other shapes and sizes are present in many mammals including some larger notoungulates (e.g., the toxodontid Nesodon; Scott 1912). The relative importance of function (versus phylogeny) in the presence of this structure in Protypotherium cannot be inferred with confidence.
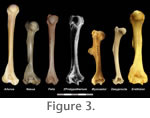
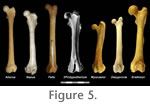
In overall form the humerus is most similar to that of Felis (Figure 3), here considered a generalist. The most significant differences between the two include: slightly more prominent greater tubercle in Felis; more proximally extended (and laterally arched) lesser tubercle in Protypotherium; longer supinator crest in Felis; and larger medial epicondyle in Protypotherium. The humerus of Protypotherium lacks the extreme extension of the greater tubercle seen in artiodactyls. It also lacks the large, distally positioned deltoid tuberosity and broad supinator ridge commonly seen in arboreal and semifossorial mammals. Radius and Ulna (Figure 2)In contrast to the humerus, the ulna and radius of Protypotherium are more similar to those of a semifossorial mammal such as Myocastor than to a generalized mammal like Felis. Both elements are proportionately shorter and more robust than those of Felis, and the ulna bears a large, medially curved olecranon with an anterosuperiorly oriented fossa for muscle attachment present along its medial surface. As in Felis and more cursorial mammals, however, the radial notch of the ulna is pronounced, suggesting relatively little movement of the radial head.
Several features of the femur suggest cursorial adaptations in the hind limb. The greater trochanter, which extends proximally above the level of the head, would have reduced the mobility of the hip joint and created a greater lever arm for the gluteal hip extensors. The short, robust femoral neck also would have reduced hip mobility.
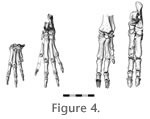
Despite the cursorial features of the femur, it is relatively short and robust compared to those of small artiodactyls and carnivorans such as Felis and Canis. In this regard, it is more similar to semifossorial rodents (e.g., Myocastor, Cuniculus). The third trochanter is more distinct in Protypotherium than in extant cursorial mammals of similar size, but is much more proximally positioned than in some arboreal mammals (e.g., Erethizon); in other arboreal mammals (e.g., Ailurus) the third trochanter is essentially absent. Given the presence of a prominent third trochanter in most perissodactyls (regardless of size or locomotor habit), this trait could be a phylogenetic correlate rather than a functional one in Protypotherium and other notoungulates. The patellar groove is intermediate in breadth between cursorial mammals and arboreal/semifossorial ones. Tibia and Fibula (Figure 2)The prominence of the cnemial crest in Protypotherium is more similar to the condition in semifossorial rodents such as Myocastor and Cuniculus than cursorial carnivorans and ungulates. The relative size of the fibula is also similar, as is the presence of a variable degree of distal fusion (either syndesmosis or synostosis) between the tibia and fibula (though the proximal fusion in rodents is opposite the distal fusion of Protypotherium). Both proximal and distal tibiofibular fusion also occurs in bounding/cursorial mammals (e.g., lagomorphs), as well as hegetotheriids, a group of typothere notoungulates generally regarded as cursorial (Cifelli 1985; Elissamburu 2004; Croft and Anaya 2006). Proximal tibiofibular fusion in Protypotherium therefore could be functionally related either to strengthening the hind limbs to brace the animal during digging or to limiting movement between these elements during bounding locomotion. Pes (Figure 4)Like the manus, the pes suggests Protypotherium was not a particularly proficient digger or climber. The astragalar trochlea is high and narrow, indicating movements limited to the parasagittal plane at the crurotarsal joint. Arboreal and semifossorial mammals typically have a lower, broader astragalar trochlea permitting greater mobility. Similarly, both the neck of the astragalus and the calcaneal tuber are elongate, more typical of the distal limb elongation seen in cursorial mammals. The absence of the first digit is compatible with many locomotor habits. |
|