Classification and Distribution of
South Atlantic Recent Polycystine Radiolaria
Plain-Language Summary:
 Radiolarians are exclusively marine,
planktonic (living in suspension in the water column),
single-celled organisms (protists). They normally dwell in
open-ocean waters, and are absent or rare near the coast. As most
other closely related planktonic protists, like the Foraminifera
(Figure 1) and the Acantharia (Figure
2), radiolarians are
omnivorous particle feeders which catch their prey (small algal
cells, other protists, small crustaceans, etc.) with the aid of
their sticky cytoplasmic strands protruding from the cell body,
the pseudopodia (axopodia - thicker, more rigid and provided with
a central shaft, and the filopodia - thinner and more flexible
than the former, article: Figure 1A).
Radiolarians are exclusively marine,
planktonic (living in suspension in the water column),
single-celled organisms (protists). They normally dwell in
open-ocean waters, and are absent or rare near the coast. As most
other closely related planktonic protists, like the Foraminifera
(Figure 1) and the Acantharia (Figure
2), radiolarians are
omnivorous particle feeders which catch their prey (small algal
cells, other protists, small crustaceans, etc.) with the aid of
their sticky cytoplasmic strands protruding from the cell body,
the pseudopodia (axopodia - thicker, more rigid and provided with
a central shaft, and the filopodia - thinner and more flexible
than the former, article: Figure 1A). 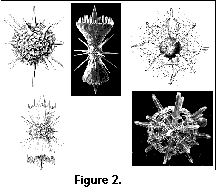 In addition, many species host living algal
cells in their cytoplasm (Figure 3); the animal benefits from the organic
matter produced photosynthetically by the symbiont alga, while
the alga takes advantage of the various substances which the
animal produces in its metabolism and would otherwise have to
discard in the water. Most radiolarian species are solitary, each
individual being represented by a single cell, yet a few species
form colonies in which many cells, enclosed in a gelatinous
cylindrical or otherwise shaped sheath (article: Figs. 1C, 15.6, 15.7, 15.8, 15.11, 15.12),
are connected with each other by cytoplasmic strands.
Radiolarians (and the closely related Acantharia, Figure 2)
differ from other protists in having their cytoplasm divided into
2 regions by a proteic perforated membrane - the central capsule
membrane.
In addition, many species host living algal
cells in their cytoplasm (Figure 3); the animal benefits from the organic
matter produced photosynthetically by the symbiont alga, while
the alga takes advantage of the various substances which the
animal produces in its metabolism and would otherwise have to
discard in the water. Most radiolarian species are solitary, each
individual being represented by a single cell, yet a few species
form colonies in which many cells, enclosed in a gelatinous
cylindrical or otherwise shaped sheath (article: Figs. 1C, 15.6, 15.7, 15.8, 15.11, 15.12),
are connected with each other by cytoplasmic strands.
Radiolarians (and the closely related Acantharia, Figure 2)
differ from other protists in having their cytoplasm divided into
2 regions by a proteic perforated membrane - the central capsule
membrane. 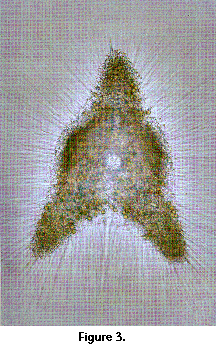 This membrane separates the inner part of
the cytoplasm, known as endoplasm, which contains the nucleus or
nuclei of the cell, from the outer part, the ectoplasm (article: Figure
1A). With but a few
exceptions, the cytoplasm encloses a shell or skeleton made up of
silica. Radiolarian skeletons are extremely diverse, and they are
the key feature on the basis of which this subclass is classified
into smaller units (classes, superorders, orders, families,
genera and species, Figure 4).
This membrane separates the inner part of
the cytoplasm, known as endoplasm, which contains the nucleus or
nuclei of the cell, from the outer part, the ectoplasm (article: Figure
1A). With but a few
exceptions, the cytoplasm encloses a shell or skeleton made up of
silica. Radiolarian skeletons are extremely diverse, and they are
the key feature on the basis of which this subclass is classified
into smaller units (classes, superorders, orders, families,
genera and species, Figure 4).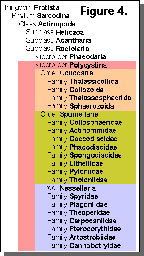 In the simplest case the skeleton is
represented by several isolated straight or branching spines
loosely distributed in the cytoplasm (article: Figure
15.4). In most species,
however, more or less elaborate structures develop, like one or
more concentric spherical shells with protruding spines or tubes
(article: Figs. 15.20,
15.26,
15.40,
15.47,
15.48),
lentil-like disks (article: Figs. 15.62, 15.63),
three-armed, propeller-like, structures (article: Figs. 15.70, 15.71),
spiral meshworks (article: Figs. 15.86, 15.87,
15.88),
elongated constructions with one or several inflated sections or
segments (article: Figs. 15.136, 15.137,
15.141,
15.163),
widely open funnel-like shells resembling a Chinese peasant hat
(article: Figure 15.148), etc. In all cases, the skeleton is
composed of siliceous bars or rods which merge with each other
leaving empty spaces between them - the pores (article: Figure
2F). A brief review of
polycystine biology, ecology, feeding and reproduction is
presented in this paper.
In the simplest case the skeleton is
represented by several isolated straight or branching spines
loosely distributed in the cytoplasm (article: Figure
15.4). In most species,
however, more or less elaborate structures develop, like one or
more concentric spherical shells with protruding spines or tubes
(article: Figs. 15.20,
15.26,
15.40,
15.47,
15.48),
lentil-like disks (article: Figs. 15.62, 15.63),
three-armed, propeller-like, structures (article: Figs. 15.70, 15.71),
spiral meshworks (article: Figs. 15.86, 15.87,
15.88),
elongated constructions with one or several inflated sections or
segments (article: Figs. 15.136, 15.137,
15.141,
15.163),
widely open funnel-like shells resembling a Chinese peasant hat
(article: Figure 15.148), etc. In all cases, the skeleton is
composed of siliceous bars or rods which merge with each other
leaving empty spaces between them - the pores (article: Figure
2F). A brief review of
polycystine biology, ecology, feeding and reproduction is
presented in this paper.
Radiolarians inhabit all major
oceans and all depths of the water column, down to over
8,000-10,000 m. Different species are usually more or less
loosely associated with different depth ranges, yet the greatest
majority prefer the uppermost layers, with highest cell densities
and specific diversities occurring between 5-10 and approximately
100-150 m (article: Figure 12A, B). This work summarizes current knowledge on
the geographic and vertical distribution of the 164 living
polycystine morphotypes (species-groups, species, and subspecific
categories) so far recorded or presumed to occur in the South
Atlantic Ocean (0 to 60°S, from the South American coasts to the
coasts of Africa). However, because the area concerned covers
from equatorial to Antarctic waters, and since polycystine
radiolarians tend to inhabit all oceans within the same climatic
zone, the review covers most of the common species worldwide.
 Two major groups, or superorders, are
recognized within the Radiolaria: the Polycystina and the
Phaeodaria (Figure 4). Among other traits, the two differ in the
chemical composition of their skeletons: while in the Polycystina
these are made up of amorphous silica (basically, the composition
of sand grains), in the Phaeodaria (Figure 5) the silica is supported by an organic
matrix.
Two major groups, or superorders, are
recognized within the Radiolaria: the Polycystina and the
Phaeodaria (Figure 4). Among other traits, the two differ in the
chemical composition of their skeletons: while in the Polycystina
these are made up of amorphous silica (basically, the composition
of sand grains), in the Phaeodaria (Figure 5) the silica is supported by an organic
matrix. 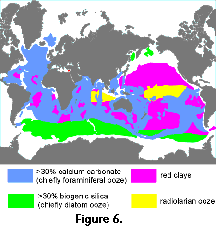 Representatives of both groups sink to the
ocean floor upon death, but differences in the composition of
their skeletons are responsible for the fact that while
polycystine skeletons can withstand dissolution during and after
descent, phaeodarian shells usually dissolve even before reaching
the bottom. Thus, large areas of the ocean floor are covered with
a thick layer of polycystine shells, also known as radiolarian
ooze (Figure 6). The Superorder Polycystina comprises 3
orders and 20 families (Figure 4). This paper contains
descriptions of these taxonomic categories, as well as brief
diagnoses and illustrations of the 164 known South Atlantic
morphotypes. A profusely illustrated glossary aimed at the less experienced student
defines the terms and morphological details useful for diagnostic
purposes.
Representatives of both groups sink to the
ocean floor upon death, but differences in the composition of
their skeletons are responsible for the fact that while
polycystine skeletons can withstand dissolution during and after
descent, phaeodarian shells usually dissolve even before reaching
the bottom. Thus, large areas of the ocean floor are covered with
a thick layer of polycystine shells, also known as radiolarian
ooze (Figure 6). The Superorder Polycystina comprises 3
orders and 20 families (Figure 4). This paper contains
descriptions of these taxonomic categories, as well as brief
diagnoses and illustrations of the 164 known South Atlantic
morphotypes. A profusely illustrated glossary aimed at the less experienced student
defines the terms and morphological details useful for diagnostic
purposes.
 The fact that polycystine radiolarians are
preserved in the sediments after death has been instrumental in
their application to paleontological studies. The earliest
records of their shells in fossiliferous deposits date back to
the Cambrian (1,000 million years before the present).
Radiolarian remains in the sediments allow a variety of studies,
including determinations of the age of the sediments that contain
them, analyses of the spatial relationships between sedimentary
layers (of particular importance for the oil industry), studies
of the geologic evolution of the continental land masses and
ocean basins, etc. Investigation of the changes in the morphology
of radiolarian skeletons through time yields first hand
information on the evolution of the group, and on the rates and
modes of species formation and extinction (Figure
7). During the last decades
particular attention has been dedicated to the analysis of
climatic changes in various geographic areas. These studies have
been partly fueled by concerns associated with global warming and
the greenhouse effect. Because the observed changes in the
climatic patterns of the last couple of centuries are most
probably associated both with man-induced modifications of the
atmosphere, and with "natural" climatic cycles (chiefly
related with changes in the parameters which define the
earth’s rotation around the sun), understanding of these
cycles in the past centuries to millennia, the object of
paleoecology and paleoclimatology, is crucial for interpreting
today's climatic changes and forecasting future trends.
Polycystine radiolarians are among the very few tools available
for investigating these changes in the oceanic realm. The
techniques in question are based on comparisons of the
radiolarian assemblages in the water column and/or in the topmost
sedimentary layer (which are assumed to represent more or less
faithfully the living populations), with those retrieved from
sediments buried deeper under the sea floor (and therefore older
in age). Each assemblage is characterized by a particular
combination of species in more or less defined proportions.
Because radiolarians (as most other organisms) are sensitive to
environmental conditions, different species and species
proportions in the assemblages are indicative of different living
conditions, in particular water temperature and productivity.
Thus, qualitative and quantitative differences between a Recent
assemblage and one produced, for example, 50 years ago, can offer
valuable information on dissimilarities in the prevailing
environmental conditions that engendered the observed changes in
the radiolarian assemblages.
The fact that polycystine radiolarians are
preserved in the sediments after death has been instrumental in
their application to paleontological studies. The earliest
records of their shells in fossiliferous deposits date back to
the Cambrian (1,000 million years before the present).
Radiolarian remains in the sediments allow a variety of studies,
including determinations of the age of the sediments that contain
them, analyses of the spatial relationships between sedimentary
layers (of particular importance for the oil industry), studies
of the geologic evolution of the continental land masses and
ocean basins, etc. Investigation of the changes in the morphology
of radiolarian skeletons through time yields first hand
information on the evolution of the group, and on the rates and
modes of species formation and extinction (Figure
7). During the last decades
particular attention has been dedicated to the analysis of
climatic changes in various geographic areas. These studies have
been partly fueled by concerns associated with global warming and
the greenhouse effect. Because the observed changes in the
climatic patterns of the last couple of centuries are most
probably associated both with man-induced modifications of the
atmosphere, and with "natural" climatic cycles (chiefly
related with changes in the parameters which define the
earth’s rotation around the sun), understanding of these
cycles in the past centuries to millennia, the object of
paleoecology and paleoclimatology, is crucial for interpreting
today's climatic changes and forecasting future trends.
Polycystine radiolarians are among the very few tools available
for investigating these changes in the oceanic realm. The
techniques in question are based on comparisons of the
radiolarian assemblages in the water column and/or in the topmost
sedimentary layer (which are assumed to represent more or less
faithfully the living populations), with those retrieved from
sediments buried deeper under the sea floor (and therefore older
in age). Each assemblage is characterized by a particular
combination of species in more or less defined proportions.
Because radiolarians (as most other organisms) are sensitive to
environmental conditions, different species and species
proportions in the assemblages are indicative of different living
conditions, in particular water temperature and productivity.
Thus, qualitative and quantitative differences between a Recent
assemblage and one produced, for example, 50 years ago, can offer
valuable information on dissimilarities in the prevailing
environmental conditions that engendered the observed changes in
the radiolarian assemblages.
These techniques, also used widely
with several other marine shelled organisms, rely heavily on our
capacity to differentiate between species, as well as on adequate
knowledge of their environmental preferences. Although the
examples above may give the impression that different radiolarian
species differ widely in their morphology, in reality there are
at least as many "confusing" species as clearly
identifiable ones. In his 1887 cornerstone monograph on these
organisms, the German naturalist Ernst Haeckel described some
3700 polycystine species; today it is widely recognized that a
very significant proportion of these "species" are
nothing but developmental stages, poorly preserved (partly
dissolved) organisms, slightly different morphotypes of the same
species, etc. Unfortunately, misidentifications and establishment
of spurious species (that is, "species" which are
synonymous with others, already described previously) is a common
phenomenon in radiolarian literature. 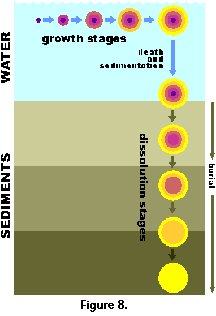 This is partly explained by the rather
intricate morphology of many radiolarians, requiring careful
observation and very good knowledge of the group. In addition,
different developmental stages of the same organism often differ
in features that are traditionally considered as specific or even
generic traits. Thus, a species whose adults possess 5 concentric
spherical shells starts as a single tiny sphere with large pores,
then the second sphere is formed, then the third, and so on
(article: Figure 14). In Haeckel’s classification scheme,
all these growth stages would qualify as different species.
Furthermore, when this shell drops to the ocean floor and starts
dissolving slowly, the innermost spheres, with thinner walls,
disappear first, thus yielding a new set of different
"species" (Figure 8).
This is partly explained by the rather
intricate morphology of many radiolarians, requiring careful
observation and very good knowledge of the group. In addition,
different developmental stages of the same organism often differ
in features that are traditionally considered as specific or even
generic traits. Thus, a species whose adults possess 5 concentric
spherical shells starts as a single tiny sphere with large pores,
then the second sphere is formed, then the third, and so on
(article: Figure 14). In Haeckel’s classification scheme,
all these growth stages would qualify as different species.
Furthermore, when this shell drops to the ocean floor and starts
dissolving slowly, the innermost spheres, with thinner walls,
disappear first, thus yielding a new set of different
"species" (Figure 8).
As indicated above,
species-specific distributional data are of primary importance
for using the polycystines for paleoclimatic analyses. These data
are derived from (1) collections of living radiolarians at
various depths in the plankton; (2) collections of radiolarian
shells settling towards the sea floor by means of sediment traps
- variously shaped containers which intercept the rain of
sedimenting particles from the upper water layers at depth; and
(3) collections of samples of surface and subsurface sediments,
where radiolarian shells accumulate after death. Information
conveyed by these different sources may differ significantly.
Thus, seasonal cycles and vertical distribution patters, among
others, may strongly bias interpretations based on planktonic
samples. Assemblages recovered in sediment traps and bottom
sediments, in turn, may differ from the corresponding planktonic
populations due to destruction of the shells by grazers,
horizontal displacement of the settling shells on their way to
the sea floor by intermediate and deep currents, partial or total
dissolution of some fragile species during and after
sedimentation, etc. (article: Figure 6). The mechanisms involved and the
concomitant potential biases for paleoclimatic interpretations
are discussed in detail in the article.
Demetrio Boltovskoy, Departamento de
Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, 1428, Buenos Aires, Museo Argentino
de Ciencias Naturales "Bernardino Rivadavia", and
CONICET, Argentina