Interpreted three-dimensional morphology of Phycosiphon incertum
The trace fossil Phycosiphon was first described by
Fisher-Ooster (1858) from Gurnigel Flysch strata of Maastrichtian age (van Stuijvenberg 1979) in the western part of Switzerland (see Wetzel and Bromley 1994). Study of topotype material facilitated the proposition of an emended diagnosis as follows: "Extensive small-scale spreite trace fossils comprising repeated narrow, U-shaped lobes enclosing a spreite in millimetre to centimetre scale, branching regularly or irregularly from an axial spreite of similar width. Lobes are protrusive, mainly parallel to bedding/seafloor. However, the plane enclosing their width may lie horizontally, obliquely or even vertically to bedding/seafloor." (Wetzel and Bromley 1994, p. 1400).
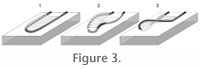 In emending the diagnosis, the authors allowed for a strong vertical component to the fecal string, which is not evident in the type material. A vertical or oblique looped fecal string is present in other similar material collected from modern depositional settings (Wetzel and Wijayananda 1990;
Wetzel and Bromley 1994). The diagnostic spreite have not, however, been fully documented from such material. The re-description of the type material by
Wetzel and Bromley (1994) included review of Anconichnus
Kern 1978, recognizing the latter as junior synonym of their emended Phycosiphon (i.e., Anconichnus is interpreted to be a morphotype of Phycosiphon with oblique to vertically oriented spreiten-bearing limbs). Supplementary block diagram models for Phycosiphon are needed to encompass cross sections of non-bedding-parallel burrows (Figure 3). In emending the diagnosis, the authors allowed for a strong vertical component to the fecal string, which is not evident in the type material. A vertical or oblique looped fecal string is present in other similar material collected from modern depositional settings (Wetzel and Wijayananda 1990;
Wetzel and Bromley 1994). The diagnostic spreite have not, however, been fully documented from such material. The re-description of the type material by
Wetzel and Bromley (1994) included review of Anconichnus
Kern 1978, recognizing the latter as junior synonym of their emended Phycosiphon (i.e., Anconichnus is interpreted to be a morphotype of Phycosiphon with oblique to vertically oriented spreiten-bearing limbs). Supplementary block diagram models for Phycosiphon are needed to encompass cross sections of non-bedding-parallel burrows (Figure 3).
The Mud-Filled "Marginal Burrow"
The most visually striking part of Phycosiphon, and all phycosiphoniform burrows in cross section, is the marginal burrow, which is generally filled with dark clay-grade material, is usually less than 1mm in diameter and is surrounded by a silty halo. The marginal burrow has not been demonstrated to self-cross (Bromley 1996). The marginal burrow of any given lobe of Phycosiphon sensu lato may be in any orientation relative to bedding (Kern 1978;
Wetzel and Wijayananda 1990;
Wetzel and Bromley 1994;
Bromley 1996;
Figure 3). Detailed three- dimensional imaging of the marginal burrow of a Phycosiphoniform burrow has been undertaken recently (Naruse and Nifuku 2008), demonstrating that the sub-horizontal to oblique limbs may lie above one another. Neither a siltstone halo nor spreiten were reconstructed, perhaps because of a lack of lithological contrast. These burrows have been assigned to Phycosiphon incertum (Naruse and Nifuku 2008), though we consider that the lack of a full complement of ichnotaxobases precludes confident ichnotaxonomic assignment of this material.
 Existing models for the orientation of lobes in Phycosiphon incertum (Wetzel and Bromley 1994) suggest that: 1) oblique lobes are most common in sandstone; 2) the same taxon in laminated siltstones and mudstones produces bedding-parallel lobes [comparable to the type material]; and 3) that lobes in homogeneous silty mudstones are commonly randomly oriented. Existing models for the orientation of lobes in Phycosiphon incertum (Wetzel and Bromley 1994) suggest that: 1) oblique lobes are most common in sandstone; 2) the same taxon in laminated siltstones and mudstones produces bedding-parallel lobes [comparable to the type material]; and 3) that lobes in homogeneous silty mudstones are commonly randomly oriented.
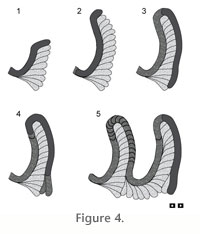
The marginal tube of Phycosiphon is looped, and defines the outer margin of spreiten-bearing regions that are discussed in detail below. A series of these curved probes are developed on one margin of the trace fossil in bedding-parallel material (Figure 4). The tube, which is surrounded by very thin "mantle" of coarser grained sediment (from which the original clay-grade material has been removed by the activity of the trace maker), is generally considered to be composed of fecal material selectively collected by deposit feeding activity in the central spreiten-bearing region.
Spreiten and Hhalos in Phycosiphon
Spreiten are positioned inside of the marginal tube and are considered to consist of zones of sediment that have been processed during feeding. The outer curves of the spreite are orientated in the direction of progressive feeding (e.g.,
Wetzel 1983;
Wetzel and Bromley 1994;
Bromley 1996;
Seilacher 2007;
Figure 4 including animation). It is anticipated that individual spreite would be meniscate if vertically sectioned through the axis of a lobe. Such a cross section has never been figured, perhaps due to either a lack of lithological contrast between spreite or the small size of most Phycosiphon.
In some cases the mud-filled tube and mantle are not associated with a spreiten bearing loop. This phenomenon was attributed to locomotory behaviour by the trace-making organism in its search for a new region of rich organic detritus (Wetzel and Bromley 1994). It is implied that when an organic-rich area is found by the trace maker, that the full spreiten-forming behaviour would resume.
The preservation of spreiten and mantle is highly dependent upon sufficient grain size contrast in the bioturbated sediment. If there is no variability in grain size in the host sediment there is little potential for spreiten formation. It has also been considered that spreiten are best preserved at sand-mud interfaces (Fu 1991). The clay-rich marginal tube is commonly the most prominent feature seen in field material. Some degree of mantle and spreiten preservation is generally seen in cross-section.
The marginal tube is generally filled with dark coloured clay-grade material and is surrounded by a thin, pale mantle of coarser grains, lithologically similar to the spreiten (Wetzel and Bromley 1994). The combination of pale mantle and spreiten material around the dark mudstone core gives rise to the colloquial term "frogspawn texture" (Bromley 1996;
Figure 1).
|