|
|
REsults
Virtual skeletal mount of Plateosaurus
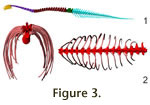 Vertebral column. Plateosaurus has 10 cervicals (plus a
rudimentary proatlas), 15 dorsal and three sacral vertebrae. The tail is
composed of 45 vertebrae in GPIT1. Figure 3.1 shows a lateral view of the
cervical, dorsal, and caudal series of GPIT1 in osteologically neutral position. Vertebral column. Plateosaurus has 10 cervicals (plus a
rudimentary proatlas), 15 dorsal and three sacral vertebrae. The tail is
composed of 45 vertebrae in GPIT1. Figure 3.1 shows a lateral view of the
cervical, dorsal, and caudal series of GPIT1 in osteologically neutral position.
The cervical column articulates in a nearly straight line, parallel to the long
axis of the cervical centra. However, this line is not perpendicular to the
anterior and posterior faces of the vertebrae, but ascends anteriorly.
Therefore, ONP displaces the head dorsally, resulting in a skull position at or
above shoulder height, depending on the angle of the anterior face of the first
dorsal relative to the exterior.
The anterior dorsals arch dorsally (dorsals 1 through 4), while the middle part curves
markedly ventrally (dorsals 5 through 10). Further posteriorly the dorsal column is
nearly straight. In all, the downward curve results in a ventral rotation of the
long axis of dorsal 5 by 22°, while the anterior upward curve angles the base of the
neck up by 12° compared to dorsal 5, and 15° down compared to D15. Therefore, if the
sacrum is placed with its long axis horizontal, the neck is attached at a slight
downward angle, and the 'stepping up' of the cervical is required to bring the
head to above shoulder level in ONP. This position is similar to that of SMNS
13200 as figured in Huene (1926). The slightly different curvature of SMNS 13200
can be explained by the damaged dorsal 6 and the slight disarticulation between
dorsals 4 and 5 (Huene 1926: plate II).
In the anterior tail, the neutral pose placing the centra faces parallel (ONP)
cannot be created. Nearly all vertebra show slight keystoning. As noted by
Moser
(2003), this keystoning is intrinsic to the osteology, but its degree varies. In
GPIT1 the variance is lower than in the material from Ellingen (Germany)
described in detail by Moser
(2003), which may be caused by a lower degree of taphonomic deformation. If the centra are placed at an angle to bring their
anterior and posterior faces into a parallel position, the haemal arches can not
be fitted to their articulation surfaces on the vertebrae. Neutral pose is
supposed to provide the maximum contact of articular surfaces. Omitting the
haemal arches, or moving them out of close articulation, would violate that
principle. To accommodate the zygapophyses in articulation dorsally and the
haemal arches in close articulation ventrally, the intervertebral disks were
reconstructed as having slightly greater thickness ventrally than dorsally,
i.e., slightly wedge-shaped. Adjustment for the haemal arches results in a
straight tail, with varying intervertebral disc thickness and shape.
Ribcage. The articulations between ribs and dorsal
vertebrae show that the anterior body was narrow from side to side (Figure 3.2).
As in all dinosaurs, the ribs of Plateosaurus have two heads (Huene
1926), the dorsal capitulum that articulates with the diapophysis on the
transverse process of the vertebra, and the ventral tuberculum, which contacts
the parapophysis. The ribs move by rotating around the axis connecting the two
articulations (Mallison in press). Whether the rib motion enlarges the ribcage
volume, e.g., for breathing in as in humans, or whether the rib motion has no
influence on the body volume depends on the orientation of the axis. In
Plateosaurus, the first five dorsal ribs move antero-posteriorly, while all
later body ribs swing outwards, increasing the volume of the body cavity (Mallison
in press). For the analysis presented here, an intermediate position of the ribs
was chosen. In- or exhalation does not significantly change the position of the
COM, because there is practically no antero-posterior motion of large tissue
volumes.
Pectoral girdle and forelimb. The pectoral girdle and
the functionally associated axial elements are incompletely preserved in GPIT1.
Only the co-ossified scapula and coracoid are preserved on each side, while the
left clavicle and sternals are missing. The right clavicle is partially
preserved and attached to the coracoid by sediment. It is a thin rod, as
described by Huene (1926). The scapulacoracoids follow the usual prosauropod
pattern in their general morphology, but show a number of peculiarities (Remes
2008), some of which have a profound impact on the interpretation of possible
locomotion poses. Most importantly, the glenoid is a simple trough, restricted
to the caudal margin of the scapulocoracoid. It does not extend to the lateral
surface of scapula or coracoid, and only the anteromedial rotation of the entire
scapulacoracoids leads to a caudoventrolateral orientation of the glenoid.
If the scapula is placed at an angle shallower than 45° in reference to the
sacrum1st dorsal line, either the pectoral girdle projects forward
beyond the second to last cervical, a position contrary to all articulated
finds, or the scapular blade overlaps beyond the fifth dorsal. Due to the
lateral expansion of the ribcage during breathing, this would lead to lateral
displacement of the dorsal end of the scapula, and thus require a hinge-joint
motion between the coracoids. The forelimbs would therefore move laterally with
each breath. Additionally, any force pressing dorsally on the glenoid, e.g.,
compressive forces in the forelimb during locomotion, would tend to rotate the
scapula anterodorsally. A placement of the scapula steeper than 65° is
unreasonable, as it would lead to a nearly horizontal orientation of the
coracoids, and direct the glenoid exclusively posteriorly.
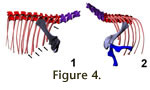 Angles of the scapula between 45° and 65° appear reasonable, as they do not push
the coracoids too far forward, but also place the tip of the scapular blade at
the level of the fourth dorsal (Figure 4.1). Such a position, with tightly
spaced coracoids, is biomechanically advantageous, as the girdle then can form a
strong brace, both for projecting large forces through the arms, and as a
support structure when the animal is lying on the ground.
Remes (2008) also
suggested this arrangement based on an analysis of the myology in basal
sauropodomorphs. In Massospondylus, a close relative of Plateosaurus,
Yates and Vasconcelos (2005) found articulated clavicles that were arranged in a furcula-like manner, touching at the midline. This further confirms the narrow
arrangement of the shoulder girdle. For Plateosaurus, this condition of
touching but not co-ossified coracoids and clavicles was already explicitly
mentioned by Huene (1926) in several skeletons. The scapulae cannot be separated
laterally and shifted dorsoposteriorlys on the side of the ribcage as suggested
by the mounts in the SMNS. It is important to note that dorsolaterally shifting
the scapulocoracoids forces a quadrupedal Plateosaurus into a sprawled
forelimb posture, with an extremely reduced functional forelimb length. Angles of the scapula between 45° and 65° appear reasonable, as they do not push
the coracoids too far forward, but also place the tip of the scapular blade at
the level of the fourth dorsal (Figure 4.1). Such a position, with tightly
spaced coracoids, is biomechanically advantageous, as the girdle then can form a
strong brace, both for projecting large forces through the arms, and as a
support structure when the animal is lying on the ground.
Remes (2008) also
suggested this arrangement based on an analysis of the myology in basal
sauropodomorphs. In Massospondylus, a close relative of Plateosaurus,
Yates and Vasconcelos (2005) found articulated clavicles that were arranged in a furcula-like manner, touching at the midline. This further confirms the narrow
arrangement of the shoulder girdle. For Plateosaurus, this condition of
touching but not co-ossified coracoids and clavicles was already explicitly
mentioned by Huene (1926) in several skeletons. The scapulae cannot be separated
laterally and shifted dorsoposteriorlys on the side of the ribcage as suggested
by the mounts in the SMNS. It is important to note that dorsolaterally shifting
the scapulocoracoids forces a quadrupedal Plateosaurus into a sprawled
forelimb posture, with an extremely reduced functional forelimb length.
The width of the shoulder girdle has been differently reconstructed with the
'old' museum mounts in the SMNS separating the coracoids by more than the length
of a coracoid and placing the scapula blade almost parallel to the dorsal
series. However, these mounts show a grossly exaggerated width of the ribcage.
GPIT1 and GPIT2 were mounted with a narrower arrangement by Huene, as was AMNH
6810, another excellently preserved skeleton from Trossingen. As shown by
Mallison (2007,
in press), the ribcage is high-oval in the shoulder area, and
only widens further posteriorly. Also, the anterior ribs sweep caudally
slightly, allowing for a narrow girdle architecture, with steeply inclined
scapula blades and medially almost contacting coracoids. Motion is restricted to
antero-posterior sweeps (Mallison in press) only in the first five dorsal ribs,
while further posteriorly there is a significant lateral expansion.
Huene (1926)
already mentioned that in SMNS 13200 these five ribs are significantly thicker
and sturdier than those of more posterior dorsals (see
Figure 4.1). They also
have a flattened cross-section. This flattening is not caused by the ribs'
attachment to the sternum, as the rough thickened distal ends indicating
attachment are found on the first eight ribs. The most plausible explanation is
that only the first five ribs had to absorb large forces transmitted through the
pectoral girdle. Recently,
Fujiwara et al. (2009) indeed found that more robust
ribs are present where the M. serratus attaches to the ribcage in quadrupeds.
The proximal end of the humerus is broad and dorso-ventrally compressed. This
indicates that a rotation of the humerus head in the glenoid, as is seen in
animals with sprawling gaits (Goslow and Jenkins 1983;
Landsmeer 1983,
1984;
Meers 2003), was not possible. Due to the simple shape of the glenoid,
protraction-extension is limited to an ~80° angle, from 55° to 135° in relation
to the long axis of the scapular blade (Figure 4.2). Therefore, if the scapula
is placed at a 35° angle from the horizontal, the humerus can not be protracted
beyond vertical. The possible amount of abduction can not be ascertained, but
any angle greater than 30° leads to an instable position, in which only limited
forces can be placed on the joint. Large forces would lead to a medial shifting
of the humeral head, as the glenoid does not contain it laterally in any way.
These findings, more details on which can be found in Mallison (in press),
confirm the results of
Bonnan and Senter (2007). Due to the medial rotation of
the glenoid axis, retraction of the humerus displaces the elbow laterally. A
purely parasagittal motion of the humerus seems not possible. The cranioventral
torsion of the distal end of the humerus relative to the proximal end is usually
45° in prosauropods (Remes 2008), but only 30° in Plateosaurus.
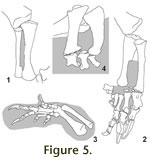 Radius and ulna can only be articulated with the distal end of the humerus if
they are placed into close proximity with each other (Mallison in press, fig.
5). The imperfect preservation of the distal humeral condyles allows separating
radius and ulna by a few millimeters or up to about 2 cm. A tight arrangement
seems more realistic, especially since the lateral side of the triangular distal
end of the ulna forms an articular surface conforming to the shape of the
proximal radius. Distally, both bones do not form contact surfaces, and are
spaced slightly apart in all articulated finds (Figure 5). In GPIT2, the left
radius and ulna each show a small deformation where they were pressed into each
other during fossilization. This relatively tight placement allows only minimal
motion between radius and ulna, making full pronation by rotation of the radius
impossible. Several other factors also indicate that Plateosaurus was not
capable of significantly pronating the hand this way: the proximal end of the
radius is oval in circumference, with a ratio between the longest and shortest
axes of 1.8:1 (Mallison in press, fig. 5). Rotating the radius head requires a
circular circumference as seen in humans and cats. Only a sliding motion, which
is not seen in any extant animal capable of pronation, remains possible. Bonnan and Senter (2007) investigated the ability to pronate the manus
using an extant phylogenetic bracket approach and found none of the extant
outgroup taxa capable of pronation. Additional evidence comes from the skeletons
that were found in articulation in Trossingen (GER) and Frick (CH). The animals
were trapped in mud (Sander 1992), their hindlimbs stuck while the forelimbs
were still free. A quadruped should in such a situation attempt to push itself
out of the mud with its forelimbs. However, not a single one of the skeletons
shows a forelimb with a pronated manus close to the body midline. Rather, the
arms are widely spread (SMNS F33), placed under the belly with the palm facing
dorsally (MSF 23 right arm), or widely abducted with strongly flexed elbow,
wrist, and fingers and medially directed palm in full supination (MSF 23 left
arm). Huene (1928: plate X) figured a quarry map of GPIT1, the only find in
which one arm potentially shows manus pronation (left forelimb). However, the
overlap pattern of the girdle and limb bones (right scapula overlaps right
humerus, left humerus wedged between the coracoids, etc.) indicates that the
skeleton was significantly displaced in a semi-macerated state, so that the
position of the manus as found is not indicative of the potential motion range
in vivo. Radius and ulna can only be articulated with the distal end of the humerus if
they are placed into close proximity with each other (Mallison in press, fig.
5). The imperfect preservation of the distal humeral condyles allows separating
radius and ulna by a few millimeters or up to about 2 cm. A tight arrangement
seems more realistic, especially since the lateral side of the triangular distal
end of the ulna forms an articular surface conforming to the shape of the
proximal radius. Distally, both bones do not form contact surfaces, and are
spaced slightly apart in all articulated finds (Figure 5). In GPIT2, the left
radius and ulna each show a small deformation where they were pressed into each
other during fossilization. This relatively tight placement allows only minimal
motion between radius and ulna, making full pronation by rotation of the radius
impossible. Several other factors also indicate that Plateosaurus was not
capable of significantly pronating the hand this way: the proximal end of the
radius is oval in circumference, with a ratio between the longest and shortest
axes of 1.8:1 (Mallison in press, fig. 5). Rotating the radius head requires a
circular circumference as seen in humans and cats. Only a sliding motion, which
is not seen in any extant animal capable of pronation, remains possible. Bonnan and Senter (2007) investigated the ability to pronate the manus
using an extant phylogenetic bracket approach and found none of the extant
outgroup taxa capable of pronation. Additional evidence comes from the skeletons
that were found in articulation in Trossingen (GER) and Frick (CH). The animals
were trapped in mud (Sander 1992), their hindlimbs stuck while the forelimbs
were still free. A quadruped should in such a situation attempt to push itself
out of the mud with its forelimbs. However, not a single one of the skeletons
shows a forelimb with a pronated manus close to the body midline. Rather, the
arms are widely spread (SMNS F33), placed under the belly with the palm facing
dorsally (MSF 23 right arm), or widely abducted with strongly flexed elbow,
wrist, and fingers and medially directed palm in full supination (MSF 23 left
arm). Huene (1928: plate X) figured a quarry map of GPIT1, the only find in
which one arm potentially shows manus pronation (left forelimb). However, the
overlap pattern of the girdle and limb bones (right scapula overlaps right
humerus, left humerus wedged between the coracoids, etc.) indicates that the
skeleton was significantly displaced in a semi-macerated state, so that the
position of the manus as found is not indicative of the potential motion range
in vivo.
The carpus is not well preserved in GPIT1 and GPIT2, but MNHB Skelett XXV from
Halberstadt has five carpals. The proximal row consists of two large elements
that are shallow triangular in plantar view. The radiale has contact with the
radius, metacarpal I, and the ulnare, as well as a small articulation with a
distal carpal. The ulnare is situated between the ulna and the distal carpals.
It is unclear whether it had any contact to the radius. The distal carpal row is
formed by a flat, box-shaped bone closely corresponding to the form of and in
close contact with the proximal end of metacarpal II, and two small rounded
elements situated between the ulna and metacarpals III and IV. These do not
block metacarpal V from contact with the ulna. The range of motion in the carpus
is difficult to determine.
Bonnan and Senter (2007) concluded that rotational or
twisting motions are made impossible by the block-like structure of the carpus.
Extension and strong flexion, however, possibly to a similar degree as in
humans, appear possible.
Metacarpals I through III articulate tightly with their proximal ends. Distally,
they show minimal splaying. Metacarpal IV also has an articulation surface for
metacarpal III, but angles laterally 20°. Metacarpal V has two distinct
articulation surfaces, and contacts both metacarpal IV and the ulna. It angles
laterally about 50° from metacarpal III.
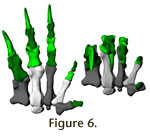 The manus digits of Plateosaurus are highly unequal, with digit I
developed as a strong grasping claw, and digit IV and V much reduced.
Paul
(1987, 1997,
2000) depicted Plateosaurus in a The manus digits of Plateosaurus are highly unequal, with digit I
developed as a strong grasping claw, and digit IV and V much reduced.
Paul
(1987, 1997,
2000) depicted Plateosaurus in a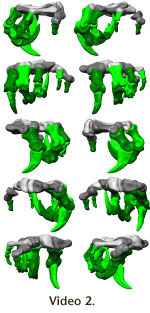 quadrupedal pose, with the
first digit medially rotated by almost 90°, so that the claw is in a horizontal
position at mid-stance. Digital manipulation of the scans of GPIT1 shows that
this angle is close to 27° during hyperextension and decreases to only 13° at
maximum flexion, because the significant size difference in the articular
condyles of metacarpal I are partially countered by an asymmetrically shaped
proximal articulation surface on the first phalanx of digit I. The significance
of the size difference lies with the larger medial condyle being subjected to
higher forces, not with a canting of the main axis of the digit. Additionally,
as pointed out by Galton (1976), in Anchisaurus, which has a similarly
shaped hand, during flexion the ungual phalanx rotates laterally, and thus
nearly lines up with the second and third digits. The latter are unremarkable,
with strong claws. However, it is important to note that hyperextension is less
pronounced than in the toes, even though there are distinct but shallow
hyperextension pits. In contrast to the first three, digits IV and V do not show
well-developed trochleae, resulting in greater freedom of motion but a reduced
ability to withstand forces. The entire hand (Figure 6,
Video 2) appears to be
adapted to strong grasping, with some ability to oppose digits IV and V. quadrupedal pose, with the
first digit medially rotated by almost 90°, so that the claw is in a horizontal
position at mid-stance. Digital manipulation of the scans of GPIT1 shows that
this angle is close to 27° during hyperextension and decreases to only 13° at
maximum flexion, because the significant size difference in the articular
condyles of metacarpal I are partially countered by an asymmetrically shaped
proximal articulation surface on the first phalanx of digit I. The significance
of the size difference lies with the larger medial condyle being subjected to
higher forces, not with a canting of the main axis of the digit. Additionally,
as pointed out by Galton (1976), in Anchisaurus, which has a similarly
shaped hand, during flexion the ungual phalanx rotates laterally, and thus
nearly lines up with the second and third digits. The latter are unremarkable,
with strong claws. However, it is important to note that hyperextension is less
pronounced than in the toes, even though there are distinct but shallow
hyperextension pits. In contrast to the first three, digits IV and V do not show
well-developed trochleae, resulting in greater freedom of motion but a reduced
ability to withstand forces. The entire hand (Figure 6,
Video 2) appears to be
adapted to strong grasping, with some ability to oppose digits IV and V.
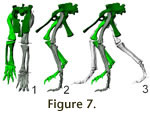
Pelvic girdle and hindlimb. The hindlimb of Plateosaurus
(Figure 7) shows a number of adaptations to cursoriality compared to basal
archosaur taxa. The femur is long, and its head is medially offset, indicating a
parasagittal limb posture, as already concluded by
Huene (1926,
1928). The
distal femoral condyles in Plateosaurus face slightly caudally, which
indicates that the knee could not be fully straightened. Combined with the
 longitudinal curvature of the femur shaft and its sub-circular cross-section
this means that Plateosaurus had a permanently flexed limb posture, and
that the femur was not held vertically during standing or at mid-stance. longitudinal curvature of the femur shaft and its sub-circular cross-section
this means that Plateosaurus had a permanently flexed limb posture, and
that the femur was not held vertically during standing or at mid-stance.
In anterior view, the probable rotation axis of the knee joint is canted against
the shaft of the femur by approximately 70° (Figure 7.1). The axis of the
proximal end is hard to determine, but appears to be roughly parallel to that of
the knee. If the femur is placed in the acetabulum so that the axis of the knee
joint is horizontal, the shaft is inclined medially and the approximate center
of the knee is placed directly ventrally from the hip joint. In
Figure 7.1, the
femur is shown adducted slightly more, so that the first pedal digit is placed
under the body midline. Such a position under or nearly under the COM is
necessary for walking gaits in bipeds, and today seen in practically all
quadrupeds.
Gatesy (1990) showed that non-avian dinosaurs use femur retraction as the major
component in caudally directed foot displacement, thus generating most of the
required force in the M. caudofemoralis. As this was certainly also the case in
Plateosaurus, it is reasonable to assume that the femur covered a large arch
during locomotion. A limit on protraction is imposed by the pubes at low
abduction angles. Retraction is not hindered by the ischia, so a maximum angle
must be estimated based on the path of the M. caudofemoralis longus and its
maximum contraction. This at best is educated guesswork, but it seems
reasonable, given the position of the tail in relation to the pelvis, to assume
that during locomotion retraction was limited to a position in which the femur
shaft is parallel to the ischia, as at this point all ischiofemoral musculature
has no retraction function left at all. If this is correct, then the angle that
the femur can cover without significant abduction is 65°.
Figure 7.2 depicts the hindlimbs and pelvic girdle in lateral view, with a suggested stride length of 1
m for a normal walk. This correlates roughly to a walking speed of 3.7 km/h,
using an average of Alexander's (1976) and
Thulborn's (1990) formulas. For this
stride length, the femora need only cover a 50° angle. For angles larger than
65°, abduction is required so that the femur passes the pubis laterally, or
retraction to beyond the level of the ischia. Compared to the neutral position
chosen here (distal end of femur directly ventral from acetabulum in anterior
view), the former requires 20° abduction. It appears therefore doubtful whether
significantly larger protractions than to the level of the pubis were possible.
Not only is all pubofemoral musculature ineffective for protraction when the
femur is close to the pubis and has a purely adducting effect. Also, there is no
greatly enlarged proacetabular process on the ilium of Plateosaurus.
Therefore, the iliofemoral muscles that still have a protracting component are
weak. Passive protraction, e.g. by transfer of rotational inertia from the shank
to the thigh, is possible, but only effective at very high limb swing speeds
(running gaits). One must assume that femur motion during normal locomotion was
limited to the mentioned 65° at medium speeds, while slower speeds probably used
less retraction. The center of mass was located in front of the acetabulum, and
mid-stance position as reconstructed here was at slight protraction (20° from
vertical, Figure 7.2).
The tibia shaft is straight, and the tibia expands slightly proximally, but
there is no distinct cnemial crest projecting anteriorly in lateral view. Knee
extension therefore did not form a major part of locomotory limb motions,
excluding the possibility of a hopping gait as suggested by
Jaekel (1913-14),
and probably also a bounding gallop as suggested by
Paul (2000). In the tibia
and fibula there is no canting between the long axis of the shaft and the
apparent joint axes. Distally, the tibia articulates tightly with the massive astragalus. Four more carpals, a calcaneum and three distal tarsals, are
preserved in GPIT1. On the left foot of MB.R.4404, three distal tarsals with
very similar shapes to those of GPIT1 are preserved, so the likeliness of
taphonomic deformation is low. The ankle is wide and flattened, with no bony
structure forming a pronounced heel, and no indication of a large cartilaginous
projection. Therefore, extension moments around the ankle were either low or
created primarily through high muscular forces. In contrast, mammals use a large
lever arm in their ankles, the tuber calcanei (Romer 1949). The ankle of GPIT1
forms a hinge joint, the axis of which is almost parallel to the knee (Figure
7.1). The range of motion is large, covering at least 170°. This would allow any
limb pose from straight to completely folded, as in resting birds. Astragali of
other individuals show a very similar morphology, if taphonomic deformation is
taken into account. As a result of the arrangement of the joint axes, placing
the femur canted inwards so that the knee axis is horizontal results in a
vertical position of the lower limb, and places the third toe ventrally below
acetabulum in anterior view (but not in lateral view, because the COM lies in
front of the acetabulum).
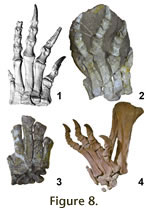 The metatarsals articulate tightly (Figure 8.1), with clearly demarcated, large
articulation facets on their lateral sides. These, however, do not run down the
shafts to a great extent, since the proximal articular ends are much wider than
the shafts. Some previous reconstructions (e.g., the 'old' SMNS mounts) have
therefore been created with splayed metatarsals. However, such a placement is
not supported by the skeletons found in Trossingen, Halberstadt, and Frick,
where all feet found in articulation show the metatarsals closely touching
proximally and almost touching distally (Figure 8.2-4). There is no splaying in
these finds, which would also be biomechanically unsound. Splayed metatarsals
result in a larger, thus heavier foot, which decreases the swing frequency of
the limb. Additionally, splaying causes lateral bending moments in the lateral
and medial metatarsals. These could result in adaptation of the shape of the
metatarsals and toes, e.g., unequal condyles on the interphalangeal joints and
asymmetrical metatarsal shaft cross sections, as well as laterally curved
metatarsal shafts, which are not present in Plateosaurus. A broader foot
would offer more lateral stability, but in extant cursors feet are reduced in
size, because the above mentioned factors outweigh the advantage of slightly
greater stability. Only animals regularly walking on instable substrates such as
snow and sand (e.g., arctic foxes and camels, respectively) show slightly larger
support areas than their relatively moving on stable ground. Even in them the
size increase in the foot is proportionally much smaller than splaying of the
metatarsals would create in Plateosaurus. The metatarsals articulate tightly (Figure 8.1), with clearly demarcated, large
articulation facets on their lateral sides. These, however, do not run down the
shafts to a great extent, since the proximal articular ends are much wider than
the shafts. Some previous reconstructions (e.g., the 'old' SMNS mounts) have
therefore been created with splayed metatarsals. However, such a placement is
not supported by the skeletons found in Trossingen, Halberstadt, and Frick,
where all feet found in articulation show the metatarsals closely touching
proximally and almost touching distally (Figure 8.2-4). There is no splaying in
these finds, which would also be biomechanically unsound. Splayed metatarsals
result in a larger, thus heavier foot, which decreases the swing frequency of
the limb. Additionally, splaying causes lateral bending moments in the lateral
and medial metatarsals. These could result in adaptation of the shape of the
metatarsals and toes, e.g., unequal condyles on the interphalangeal joints and
asymmetrical metatarsal shaft cross sections, as well as laterally curved
metatarsal shafts, which are not present in Plateosaurus. A broader foot
would offer more lateral stability, but in extant cursors feet are reduced in
size, because the above mentioned factors outweigh the advantage of slightly
greater stability. Only animals regularly walking on instable substrates such as
snow and sand (e.g., arctic foxes and camels, respectively) show slightly larger
support areas than their relatively moving on stable ground. Even in them the
size increase in the foot is proportionally much smaller than splaying of the
metatarsals would create in Plateosaurus.
All toes of GPIT1 are almost straight, in contrast to Skelett XXV (MB.R.4404)
from Halberstadt, in which the fourth toe curves laterally. Similarly, in GPIT1
toes 1 through 3 are canted slightly medially compared to the long axis of the
metatarsus, though beveled distal condyles of the metatarsals. The fourth
metatarsal, however, is canted laterally. In contrast, in Skelett XXV, toes 1
through 3 are canted strongly medially, and the base of the fourth toe is
perpendicular to the long axis of the metatarsus. It is unclear whether this
difference is caused by intraspecific variation, or indicates that the two
individuals belong to separate species.
The hindlimb digits of GPIT1 allow somewhat larger hyperextension angles than in
the forelimb, indicative of a highly digitigrade to unguligrade stance just
before toe lift-off. This adds to the greater effective limb length in the
hindlimb compared to the lower hyperextension angles in the forelimb. Further
indicators for digitigrady are the dorsoventrally flattened cross-sections of
the metatarsal shafts and the lack of proximal-distal arching in the metatarsus,
both indicating low craniocaudal bending moments.
Poses of the entire skeleton
In the following, both bipedal and quadrupedal poses are described, and their
biomechanical implications are addressed.
B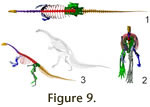 ipedal poses. For a bipedal, theropod-like posture, the
hindlimb of Plateosaurus can be placed in accordance with the evidence on
bone loading derived from longbone curvature and cross sections. The femur is
inclined forward at a protraction angle around 20°. The knee is flexed, and the
tibia and fibula inclined slightly posteriorly. The ankle is also slightly
flexed, placing the metatarsus in a vertical or, more probably, sub-vertical
position at midstance (Figures 1.2,
9.1, 9.2,
Video 1). ipedal poses. For a bipedal, theropod-like posture, the
hindlimb of Plateosaurus can be placed in accordance with the evidence on
bone loading derived from longbone curvature and cross sections. The femur is
inclined forward at a protraction angle around 20°. The knee is flexed, and the
tibia and fibula inclined slightly posteriorly. The ankle is also slightly
flexed, placing the metatarsus in a vertical or, more probably, sub-vertical
position at midstance (Figures 1.2,
9.1, 9.2,
Video 1).
Any bipedal posture is only feasible if the COM is supported by the hindfeet. If
the COM rests close to the hips, the exact orientation of the vertebral column
does not matter. If it is placed further forward, so that the hindlimb does not
support it when the back is in a subhorizontal position, the simplest way to
change the anteroposterior position of the COM is to tilt the vertebral column.
The steeper it is placed, the further back the COM moves.
Huene 1928 suggested a
fairly steep position for rapid locomotion (Figure 9.3), similar to the then
prevailing view of Iguanodon (Dollo 1893) and other bipedal dinosaurs.
Later, a paradigm shift lead to almost universal agreement that most dinosaurs
had subhorizontal backs when walking bipedally (e.g.,
Norman 1980;
Bakker 1986;
Paul 1987). The main reason for the latter posture is the far greater femur
retraction range that it allows compared to a more upright posture.
Plateosaurus can be placed bipedally in either position (Figures 1.2,
9.1-3), but the more upright posture limits locomotion speed significantly. An
upwards angle of at least 45° is required for the long axis of the sacrum to
create a significant backwards shift of the COM. However, already at 30° (Figure
9.3), the femur must be retracted to the level of the ischia, so that only very
small steps are possible. Also, if the COM lies so far posteriorly that this
pose (30° upwards rotation) is balanced, rotating the body around the hips
requires very little energy, due to the small moment arm. Therefore, it seems
more reasonable to assume that a bipedal Plateosaurus might feed with a
steeply inclined vertebral column (Figure
9.3), but use a subhorizontal posture
(Figure 1.2) for locomotion.
In a bipedal pose with a subhorizontal back, the neutral pose of the vertebral
column leads to a head position above the highest point of the back. The animal
can thus cover a 360° arc by a small neck motion alone, i.e., there is no blind
area. In order to bring the snout to the ground, e.g. for drinking, a slight
increase in hindlimb flexion and moderate ventriflexion of the anterior two
thirds of the thoracic vertebral column and maximum ventriflexion of the neck is
sufficient (Mallison in press, contra
Huene 1928), but even a slight seesaw
motion that rotates the anterior body down (as suggested to be necessary by
Huene [1928]) does not inhibit rapid flight. In contrast, to bring the hands to
the ground for grasping requires significant flexion of the hindlimbs, with
greatly increases joint torques required to sustain the pose. Essentially, the
animal must kneel or squat down when manipulating objects at ground level for a
prolonged time, or use a front limb for support. Dual-handed grasping is then
impossible. Rainforth's (2003) redescription of the ichnofossil Otozoum
pointed out that one track shows a bipedal animal, potentially a prosauropod,
using both hands, palms facing medially, to support itself, probably while
squatting down.
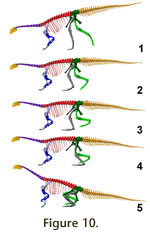 Quadrupedal poses. In a quadrupedal pose the forelimbs must be
able to take strides of a significant length. If a stride length of only 0.4 m
is to be possible, the height of the glenoid above the ground when placing the
manus on the ground can not be more than 0.6 m. Due to this extremely short
forelimb length, the vertebral column slope downwards so that the 1stdorsal
2ndsacral line slopes by 19° for a digitigrade
Figure 10.1), and 9°
for a semi-plantigrade model (Figure 10.2). In a fully plantigrade (FVP) model,
whether with a strongly protracted or nearly vertical femur position, the head
is at or slightly above the same height as the sacrum (Figure 10.3-4). These
postures bring the manus on the ground without introducing strong ventriflexion
in the dorsal series, and retain room for femur excursions during locomotion.
The effective forelimb/hindlimb length ratios (i.e., height of glenoid divided
by height of acetabulum in midstance pose) are 0.45 for the digitigrade, 0.54
for the semi-plantigrade, and 0.59 for the plantigrade model. In all these
positions the motion of the hindlimb is significantly limited compared to a
bipedal posture. In the digitigrade model, the femur can be protracted roughly
20° less than in a bipedal pose, reducing stride length. In the semi- and
plantigrade models, limb protraction requires extreme flexion of the ankle while
the free limb passes the supporting limb, due to the great length of the pes
compared to the tibia. In such a pose femur retraction cannot be the main
component of protraction unless the animal uses a sprawling posture, in direct
contradiction to the osteological evidence (see
Gatesy 1990). Quadrupedal poses. In a quadrupedal pose the forelimbs must be
able to take strides of a significant length. If a stride length of only 0.4 m
is to be possible, the height of the glenoid above the ground when placing the
manus on the ground can not be more than 0.6 m. Due to this extremely short
forelimb length, the vertebral column slope downwards so that the 1stdorsal
2ndsacral line slopes by 19° for a digitigrade
Figure 10.1), and 9°
for a semi-plantigrade model (Figure 10.2). In a fully plantigrade (FVP) model,
whether with a strongly protracted or nearly vertical femur position, the head
is at or slightly above the same height as the sacrum (Figure 10.3-4). These
postures bring the manus on the ground without introducing strong ventriflexion
in the dorsal series, and retain room for femur excursions during locomotion.
The effective forelimb/hindlimb length ratios (i.e., height of glenoid divided
by height of acetabulum in midstance pose) are 0.45 for the digitigrade, 0.54
for the semi-plantigrade, and 0.59 for the plantigrade model. In all these
positions the motion of the hindlimb is significantly limited compared to a
bipedal posture. In the digitigrade model, the femur can be protracted roughly
20° less than in a bipedal pose, reducing stride length. In the semi- and
plantigrade models, limb protraction requires extreme flexion of the ankle while
the free limb passes the supporting limb, due to the great length of the pes
compared to the tibia. In such a pose femur retraction cannot be the main
component of protraction unless the animal uses a sprawling posture, in direct
contradiction to the osteological evidence (see
Gatesy 1990).
 Subequal fore and hind limb lengths cannot be created, even with a pronated
manus and parasagittal forelimbs, as it requires femur protraction to beyond the
pubes at midstance, and to subhorizontal for limb protraction (Figure 10.5,
Video 3). Note that in this position the height of the glenoid was artificially
increased to 0.7 m by using a standing instead of a walking pose in the
forelimb. This position does not allow proper walking at all, as it limits
stride length to 0.2 m. Subequal fore and hind limb lengths cannot be created, even with a pronated
manus and parasagittal forelimbs, as it requires femur protraction to beyond the
pubes at midstance, and to subhorizontal for limb protraction (Figure 10.5,
Video 3). Note that in this position the height of the glenoid was artificially
increased to 0.7 m by using a standing instead of a walking pose in the
forelimb. This position does not allow proper walking at all, as it limits
stride length to 0.2 m.
However, all these poses were created with a pronated manus. If the forelimb is
placed in correct articulation with regards to the wrist and elbow, strong
humerus abduction is required to turn to palm ventrally. This would essentially
create a sprawling pose. However, it is not possible to rotate the humerus
around its long axis to create a subvertical antebrachium, so that the animal
would touch the ground with its ribcage. If the hindlimbs are placed in a
sprawling position (following Fraas 1912; contra
Huene 1926,
1928;
Gatesy 1990;
Christian
and Preuschoft 1996;
Christian et al. 1996), the femora must be disarticulated
from the pelvis, and the tibia and fibula must be disarticulated from the distal
femoral condyles. Alternatively to a sprawling pose, Plateosarus might
have walked on non-pronated hands, but
Bonnan and Senter (2007) show clearly
that the required adaptations are missing.
A potentially negative effect of the only quadrupedal poses in which the
hindlimbs can be moved in a realistic gait cycle (hindlimbs digitigrade,
Figure
10.1) is the forwardly inclined vertebral column combined with the low shoulder
height. At neutral articulation of the neck, the view the animal can see is
quite limited. A blind angle extends posteriorly, and lateral excursions of the
neck to 'check six' result in a large blind area on the contralateral side.
Extreme dorsiflexion of the neck is required to bring the head to a sufficient
height so that a 360° view is possible. While possibly feasible when feeding,
during rapid locomotion such a neck position carries a high risk, because all
articulations are at their bony stops. Therefore, even slight impulses, e.g.,
from stumbling, can lead to serious injury. Extant animals appear to carry their
necks close to neutral articulation during rapid locomotion (Christian and Dzemski 2007), potentially because of this risk. For Plateosaurus,
running in a quadrupedal posture carrying the neck near neutral articulation
leads to a head height below hip height, close enough to the ground that a
misstep in the forelimbs leads to an impact on the ground.
A further potential disadvantage is the maximum possible head height, and thus
size of the feeding envelope. The base of the neck is at two thirds the height
of a bipedal pose (0.99 m compared to 1.49 m). Additionally, in a bipedal pose
the animal can tilt the body up to increase shoulder and thus feeding height
(Figure 9.3). While Plateosaurus could theoretically be envisaged as an
obligate quadruped that uses quadrupedal gaits for slow locomotion, then gets up
into a bipedal pose to feed, and then back down again into a quadrupedal stance
for locomotion, this makes sense only if there are bipedal stances suitable for
feeding but not for locomotion. These could only be postures with a steeply
inclined vertebral column, akin to a rearing elephant. However, as shown above,
there are no bipedal poses that do not allow locomotion.
Body mass
The CAD model of Plateosaurus based on GPIT1 has a total volume of 740 l.
There are no indications of air sacs invading bone to form post-cervical
pneumatic cavities in Plateosaurus or other prosauropods (Wedel 2007),
but the dorsal vertebrae show shallow troughs on the centra. In combination with
the ability to breathe through rib motion, this indicates that prosauropods
probably possessed pulmonary air sacs. An extant phylogenetic bracket (EPB)
approach (Witmer 1995) confirms this: both theropods and sauropods had (and
have) bird-like lungs (Perry and Reuter 1999;
Wedel 2005,
2007;
O'Connor and Claessens 2005), as
indicated by the extensive pneumaticity of their skeletons. Birds have densities
as low as 0.73 kg/l (Hazlehurst and Rayner 1992), and sauropod density is
therefore probably best assumed to be about 0.8 kg/l or even lower (Wedel 2005).
Crocodiles have lungs that, albeit simpler in structure than those of birds, are
interpreted as a preadaptation for the formation of true air sacs and a
unidirectional lung (Perry 1998;
Farmer and Sanders 2010). Therefore, it is most
parsimonious to assume the existence of small pulmonary air sacs in
Plateosaurus, even though they did not invade bones and thus left no clear
marks on the preserved parts of the animals. Rib motion probably was the main
mode of lung ventilation in Plateosaurus, because the architecture of the
costovertebral articulations (dual headed hinge joints) is the same as is
generally the case in saurischians (basal Saurischia:
Langer 2004;Tyrannosaurus:
Hirasawa 2009), and the air exchange volume can be estimated at a value typical
for birds (Mallison in press). The air sacs may have been large, but in the
absence of solid evidence (skeletal pneumaticity) for a large size they must be
assumed to have been so small that the overall density can be modeled on that of
extant terrestrial vertebrates. As a consequence, the average density should
cautiously be
assumed at a value between 0.9 kg/l and 1.0 kg/l.
Additionally, density is not uniform across the entire animal, and various body
parts were accordingly given values that reflect the relative abundance of bone,
flesh, intestines, and air volumes in them. Skull, tail, limbs, and the pelvic
region are assumed to be heavier than water (d = 1.05 kg/l to 1.1 kg/l), while
the neck and anterior trunk region are significantly lighter (d = 0.7 kg/l).
This basic model has an average density of 0.94 kg/l and a total weight of 693
kg. Variations of the density of model parts to account for different soft
tissue amounts, details of which are given in
Table 1, result in average
densities between 0.89 kg/l and 1.13 kg/l. The basic model has a very posteriorly placed COM, so most variations were designed to move the COM
forward. This was achieved by reducing the mass of the tail and posterior trunk,
and/or increasing the mass of neck and anterior trunk. However, one variant was
produced in which the limbs were given higher density, too, although this partly
cancels the effect that a heavier neck and anterior body have on shifting the
center of mass. Total mass varies between 600 kg and 838 kg. Scaled to the size
of the largest and smallest known individuals (total length 10 m and 4.8 m,
Sander and Klein 2005) of Plateosaurus, the basic model gives a weight
range from 476 kg to roughly 4300 kg for an average density of 0.89 kg/l.
Position of the center of mass (COM)
In the basic model, the COM rests 0.23 m in front of and 0.16 m below the
acetabulum in a bipedal standing pose. The mass variations result at most in an
anterior shift by 0.06 m to 0.29 m (Table 1). The dorsoventral shift is
negligible.
Bipedal poses. In the bipedal poses created on the basis of a
best fit of the skeletal elements, in all but one the COM rests comfortably
above the support area (Figure 11.1). This is true in both the digitigrade and
the semi-plantigrade pose, and requires only modest femur protraction (maximum
value 20° from vertical, in combination with a fully digitigrade stance and a
metatarsus inclination of 65° from horizontal). For the most front-heavy mass
variation at full digitigrady the COM plots just in front of the longest digit.
Femur protraction to 25° (+5° compared to the basic model) brings the support
area under the COM in this model.
Quadrupedal poses. In all models with an inclined femur
the COM plots in the support area of the hind foot or at its anterior edge,
unless the lower hindlimb is placed at an unrealistic strong posterior
inclination (as in Figure 10.4). If the hind limb is posed as deemed
anatomically correct here, with a protracted femur, limited knee and ankle
flexion and fully digitigrade, the ratio of effective limb lengths is 0.45, and
the hindlimb carries between 90% and all of the weight. Unrealistically flexing
the knee so that the tibia is strongly inclined leads to a maximum of 35% of the
weight supported on the forelimbs, albeit with a low effective limb length ratio
of 0.52. Increased limb flexion, which results in less massively unequal limb
lengths, moves the support area forward, due to increases in femur protraction
and ankle flexion. In fact, even absurdly light-tailed versions of the model
with a significantly inflated pectoral region and neck have a COM plotting
solidly within the area of the hindfoot in midstance at effective limb length
ratios between 0.7 and 1. Subequal fore and hind limb lengths in a sprawling
position place up to 40% of the weight on the forelimbs but are osteologically
impossible.
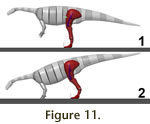 Models with a vertical femur position place more weight on the forelimbs, by
shifting the support point of the hind limbs posteriorly. However, they either
require a strongly inclined position of the lower hindlimb, which creates large
bending moments because of the long moment arm of the COM and high torques in
the knee and ankle joints, or cause an extreme difference in limb length. In
extant animals with parasagittal limbs, the hindfoot is almost always placed
under the hip joint, or close to such a position. The Plateosaurus models
with a vertical femur and strong limb flexion, however, result in a foot
position far behind the acetabulum. Such a posture causes extreme flexing
moments in the knee and ankle, and can thus be dismissed as unrealistic,
especially considering the short moment arms of the extensor muscles of the
ankle and knee. The alternative pose, with a steeply placed lower hindlimb (low
limb flexion), causes similarly low limb length ratios as those models with an
inclined femur and a slightly flexed hind limb. At most, using the unreasonable
mass distribution of the most front-heavy model (d = 1.13 kg/l), the forelimbs
carry about 41% of the body weight (Figure 11.2), with a limb length ratio of
0.45. Note, however, that for this hindlimb posture the acetabulum is not above,
but far in front of the point of support, and the hind limb thus permanently
subjected to large bending moments. Models with a vertical femur position place more weight on the forelimbs, by
shifting the support point of the hind limbs posteriorly. However, they either
require a strongly inclined position of the lower hindlimb, which creates large
bending moments because of the long moment arm of the COM and high torques in
the knee and ankle joints, or cause an extreme difference in limb length. In
extant animals with parasagittal limbs, the hindfoot is almost always placed
under the hip joint, or close to such a position. The Plateosaurus models
with a vertical femur and strong limb flexion, however, result in a foot
position far behind the acetabulum. Such a posture causes extreme flexing
moments in the knee and ankle, and can thus be dismissed as unrealistic,
especially considering the short moment arms of the extensor muscles of the
ankle and knee. The alternative pose, with a steeply placed lower hindlimb (low
limb flexion), causes similarly low limb length ratios as those models with an
inclined femur and a slightly flexed hind limb. At most, using the unreasonable
mass distribution of the most front-heavy model (d = 1.13 kg/l), the forelimbs
carry about 41% of the body weight (Figure 11.2), with a limb length ratio of
0.45. Note, however, that for this hindlimb posture the acetabulum is not above,
but far in front of the point of support, and the hind limb thus permanently
subjected to large bending moments.
In all quadrupedal poses that do not shift the pectoral girdle to an absurdly
low position on the ribcage, the COM lies significantly higher than the glenoid
(e.g., at 1.5 times the height in the model shown in
Figure 11.2). This means
that the stability of the anterior body part when supported by only one forelimb
is reduced compared to the pattern seen in nearly all extant quadrupeds, in
which the limb articulation with the trunk rests higher than or roughly at the
level of the COM.
Stability of slow gaits. The extremely posterior position of
the COM has consequences for the stability of even slow gaits. Slow locomotion
means that the body, due to its low moment of inertia, has ample time to react
to small lateral imbalances. Unless one is balanced well, much energy is
expended correcting the balance instead of moving forward. When walking slowly
and bipedally, the body must sway laterally or the feet must
be placed in front of each other, on the track midline, so that the COM passes
exactly over the support area. These two solutions, or any combination of them,
require strong limb adduction to bring the foot under the COM. A more
longitudinal support area, such as a plantigrade instead of a digitigrade foot,
or quadrupedal instead of bipedal gaits, reduce the risk of toppling as well as
the need to adduct the hindlimbs fully. A wider track is possible, creating a
wide support triangle (Henderson 2006). To be effective, the quadrupedal posture
should thus increase both the width and the length of the support area compared
to a bipedal pose. The latter, a minor factor, is obviously the case in
practically any quadruped, but the former relies on allowing a wider track in
the forelimbs and hindlimbs for creating a wide support triangle when any one
limb is lifted.
In Plateosaurus this is not the case in any osteologically possible
quadrupedal pose, even assuming manus pronation, as is best shown by a test of
the balance in a three-point support pose. If a hindfoot is lifted off the
ground in a CAE model, using the most front-heavy mass variant, in a standing
pose with only 10° femoral abduction in both limbs, the animal tends to topple
caudolaterally, because the COM lies behind the support triangle formed by the
forefeet and the remaining hindfoot. The supporting hindlimb must be adducted to
nearly the same angle as in a biped to avoid this. Therefore, a quadrupedal pose
does not offer a significant advantage with regard to stability, because an
accidental imbalance in the supporting hindlimb is as dangerous as in a bipedal
stance. A detailed discussion of walking cycles in Plateosaurus is beyond
the scope of this paper.
Potential trackways. The virtual skeleton cannot be placed in a
quadrupedal pose that allows normal locomotion. Therefore, none of the
quadrupedal trackways assigned to prosauropods, such as Tetrasauropus
unguiferus (Ellenberger 1972) or Navahopus falcipollex (Baird 1980)
were created by a plateosaurid. In contrast, the skeleton can be placed to
conform to the ichnofossil Otozoum moodii (Hitchcock 1847,
Rainforth 2003), contra
Farlow (1992).
Porchetti and Nicosia (2007) fitted the
scaled-down pes of Plateosaurus into the track of Tetrasauropus in
a plantigrade stance. As shown, this posture is highly unlikely for
Plateosaurus.
|