MODERN ANALOG
TECHNIQUE (MAT)
The Modern Analog Technique (MAT)
quantifies faunal changes within deep-sea cores in terms of
modern oceanographic conditions (Hutson 1979). The method uses a
measure of faunal dissimilarity to compare down-core samples to
each reference sample in a modern oceanographic database. Working
with late Pleistocene planktonic foraminifers, Hutson (1979) originally used cosine-theta distance to
match modern Indian Ocean samples to late Pleistocene age core
samples and then employed a weighted average of sea surface
temperature and salinity associated with the closest analogs of
each core sample to derive down-core environmental estimates. Overpeck et al. (1985) investigated the responsiveness of eight
dissimilarity coefficients to palynological changes caused by
differences in modern vegetation and applied the MAT technique to
late Quaternary pollen diagrams from eastern North America. Their
analyses suggested that whereas all coefficients give roughly
similar results, signal-to-noise measures performed better than
unweighted or equal-weight dissimilarity coefficients (Overpeck et al. 1985).
After experimentation with
different dissimilarity coefficients we chose the squared chord
distance (SCD) measure for our study:
dij =
Sigmak(pik1/2 - pjk1/2) 2 (1)
where dij
is the squared chord distance between two multivariate
samples i and j, and pik
is the proportion of species k in sample i. Squared
chord distance values can range from 0.0 to 2.0, with 0.0
indicating identical proportions of species within the samples
being compared. To illustrate the technique, a foraminifer census
from a mid-latitude North Atlantic modern (core top) sample was
compared to 223 other modern North Atlantic samples. Figure
1A 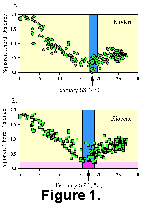 shows
that all modern samples converge on the temperature of the test
sample. Averaging the observed February SST associated with the
ten nearest analogs produces an "estimated" SST of
18.43°C. The width of the vertical blue band indicates the
standard deviation (sigma = ±1.15°C) of the values that make up
the estimated SST.
shows
that all modern samples converge on the temperature of the test
sample. Averaging the observed February SST associated with the
ten nearest analogs produces an "estimated" SST of
18.43°C. The width of the vertical blue band indicates the
standard deviation (sigma = ±1.15°C) of the values that make up
the estimated SST.
As mentioned above, this technique
has been used by others (e.g., Prell 1985, Anderson et al. 1989) to estimate late Pleistocene SST using
planktonic foraminifer census data. When working with older
(e.g., Pliocene) samples, the closest analogs are somewhat more
distant than when working with younger material (Ikeya and Cronin 1993, Dowsett 1996, Andersson 1997). This is illustrated in Figure 1B by the
horizontal pink band which indicates that no modern sample has a
squared chord distance less than 0.22 units from the Pliocene
sample selected for comparison. Nevertheless, the distribution of
points on Figure 1B clearly suggests that the most similar
samples in the modern database range between 16° and 19°C.
Averaging the 10 nearest analogs gives a temperature estimate of
17.25°C (sigma = ±1.51°C).
Another illustration of MAT is
shown in Figure 2. 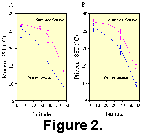 Modern
North Atlantic latitudinal SST gradients for cold and warm
seasons were developed from observed SST at four sites. Core-top
faunal data from these four sites were analyzed using MAT and the
SST estimates plotted in Figure 2A. For comparison, four Pliocene
samples representing approximately 2.795 Ma at different
latitudes were analyzed using MAT and the SST estimates were used
to draw Pliocene gradients (Fig. 2B). Whereas it is difficult to
compare the gradients based upon so few sites, it is worthwhile
to note that the Pliocene gradients are offset from the modern
gradients which are generally cooler. The Pliocene gradients are
generally less steep at low and mid-latitudes when compared to
the modern gradients. Also, seasonality (measured as summer
season minus winter season) is estimated to be greater in modern
climates than in the Pliocene. These results are reasonable and
fit well with previous studies of the Mid-Pliocene of the North
Atlantic which used independent methods of inference (Dowsett et al. 1992, Dowsett, Barron and Poore
1996). We conclude that
despite the lower levels of similarity, the MAT can provide a
powerful method for transferring modern oceanographic conditions
to Pliocene samples.
Modern
North Atlantic latitudinal SST gradients for cold and warm
seasons were developed from observed SST at four sites. Core-top
faunal data from these four sites were analyzed using MAT and the
SST estimates plotted in Figure 2A. For comparison, four Pliocene
samples representing approximately 2.795 Ma at different
latitudes were analyzed using MAT and the SST estimates were used
to draw Pliocene gradients (Fig. 2B). Whereas it is difficult to
compare the gradients based upon so few sites, it is worthwhile
to note that the Pliocene gradients are offset from the modern
gradients which are generally cooler. The Pliocene gradients are
generally less steep at low and mid-latitudes when compared to
the modern gradients. Also, seasonality (measured as summer
season minus winter season) is estimated to be greater in modern
climates than in the Pliocene. These results are reasonable and
fit well with previous studies of the Mid-Pliocene of the North
Atlantic which used independent methods of inference (Dowsett et al. 1992, Dowsett, Barron and Poore
1996). We conclude that
despite the lower levels of similarity, the MAT can provide a
powerful method for transferring modern oceanographic conditions
to Pliocene samples.
Application of this technique to
Pliocene sequences requires several additional considerations.
First, Pliocene sequences contain taxa now extinct and therefore
not in the modern database. Also, there are taxa present in the
modern database that have evolved since the Pliocene. Dowsett and Poore (1990) and Dowsett (1991) explored various ways to regroup modern
planktonic foraminifers into categories that could be used as far
back as the early Pliocene. These studies found that different
members of the genus Globigerinoides, excepting Globigerinoides
sacculifer, could be grouped into a single tropical category
with little loss of information. Likewise, at high latitudes,
members of the genus Neogloboquadrina could be grouped
together to form cold and warm end members of that genus. For
Pliocene samples, several ancestor-descendent pairs were
identified, and the Pliocene ancestors were assumed to have the
same environmental tolerances as their modern descendants. These
species groupings allowed more direct comparisons of modern and
Pliocene faunas of the North Atlantic. Similar groupings are
necessary to both modern and Pliocene data being compared by
dissimilarity measures (see below).