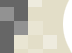Research interests
Copper homeostasis in Arabidopsis thaliana
Plants use copper (Cu) as a cofactor for a wide variety of physiological processes including photosynthesis, mitochondrial respiration, superoxide scavenging, cell wall metabolism, and ethylene sensing. In the absence of this essential micronutrient, plants display diverse symptoms, most of which affect young leaves and reproductive organs. In addition to its essentiality, Cu also limits plant growth when present in excess in contaminated soils. Cu participates in redox reactions that generate reactive oxygen species, which cause damage to lipids, proteins and nucleic acids. We are especially interested in deciphering the mechanisms of plant response to different environmental Cu statuses and its consequences in plant growth and development.
Moreover, although the temporal aspects of metal homeostasis are relatively poor studied, metals neccesity vary during the diurnal cycles of light and darkness since photosynthesis is the process with the largest metals requirements. To acquire, transport and store Cu and iron (Fe), avoid the pernicious effects of deficiency or excess, and reconcile the dynamic changes between the supply and demand, there are complex homeostasis networks which are aimed at maintaining the adequate cytosolic levels in different tissues and developmental stages.
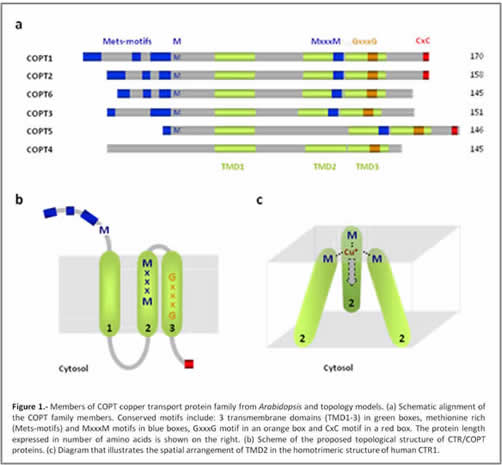
Over the past years, we have identified several essential components of the Arabidopsis thaliana Cu homeostatic network. Arabidopsis has a 6 members family of CTR-like transporters, denoted COPT in plants (Figure 1, COPT1-COPT6). Certain COPT proteins are located in the plasma membrane, where they mediate Cu uptake from the external medium. In Arabidopsis, COPT1 participates in Cu uptake from the soil through the root apex Moreover, COPT5 has been described in the membrane of the prevacuolar compartment where they are involved in the mobilization of Cu from the lumen towards the cytosol, in response to extreme Cu deficiency conditions. The mechanism of transport used by these permeases remains unknown, but the substrate is the reduced Cu+ form.
The next components in the Cu homeostasis network are the soluble cytosolic cuprochaperones, one of whose tasks is to provide Cu to the cytosolic targets, such as the CCS cuprochaperone that provides Cu to the Cu and zinc (Zn) superoxide dismutase (Cu/ZnSOD). However, other cuprochaperones, such as ATX1, deliver Cu to P-type ATPases of Cu+ (Cu+-ATPases) at the endoplasmic reticulum to pump Cu into the lumen, where it is acquired by the cuproproteins at the secretion pathway. Arabidopsis has two homologues, ATX1 and CCH, of this cuprochaperone. The Cu-transporting P-type ATPase, HMA5, is involved in Cu detoxification. We are currently investigating the function of these and additional members of the plant Cu homeostasis network and utilizing this information for applications in agriculture.
Transcriptional responses to Cu deficiency
The transcriptional response to Cu deficiency in Arabidopsis is mainly mediated by a member of the Zn finger transcription factor family named SQUAMOSA-promoter binding-like protein (SPL). SPL7 is essential for the response to Cu deficiency in vivo through its binding to GTAC motifs in the promoters of target genes (Figure 2). The repression mechanism in the presence of Cu could be mediated by the displacement of Zn by Cu+ from the Zn fingers of the transcription factor, as recently described for the Chlamydomonas reinhardtii orthologue in vitro.
One of the several strategies used by Arabidopsis when the Cu is limiting, and mediated by SPL7, consists of replacing the non-essential cuproproteins for other metaloproteins, usually Fe proteins, that carry out a similar role probably with the aim of saving Cu for essential cuproproteins, such as plastocyanin. In the case of SODs (Cu/ZnSOD and FeSOD), the substitution of the Cu form by the Fe counterpart is carried out, by the same SPL7 transcription factor under Cu limited conditions, expressing the FeSOD and degrading the Cu/ZnSOD mRNAs through the induction of the miR398 (Figure 2). Diurnal oscilations in the accumulation of the miR398 have been described which indicates the importance of the temporal factor in this response. Other Cu miRNAs are: miR397, miR408 and miR857 that are involved in the degradation of cuproproteins mRNAs as the plantacianin and laccases. Therefore it seems that regulation by miRNAs is a widespread response to Cu deficiency, as it is the case for other nutrients.